NIVELES DE ORGANIZACIÓN DE LOS SERES VIVOS · complejidad que le proporcionan unas propiedades que...
31
NIVELES DE ORGANIZACIÓN DE LOS SERES VIVOS
Transcript of NIVELES DE ORGANIZACIÓN DE LOS SERES VIVOS · complejidad que le proporcionan unas propiedades que...
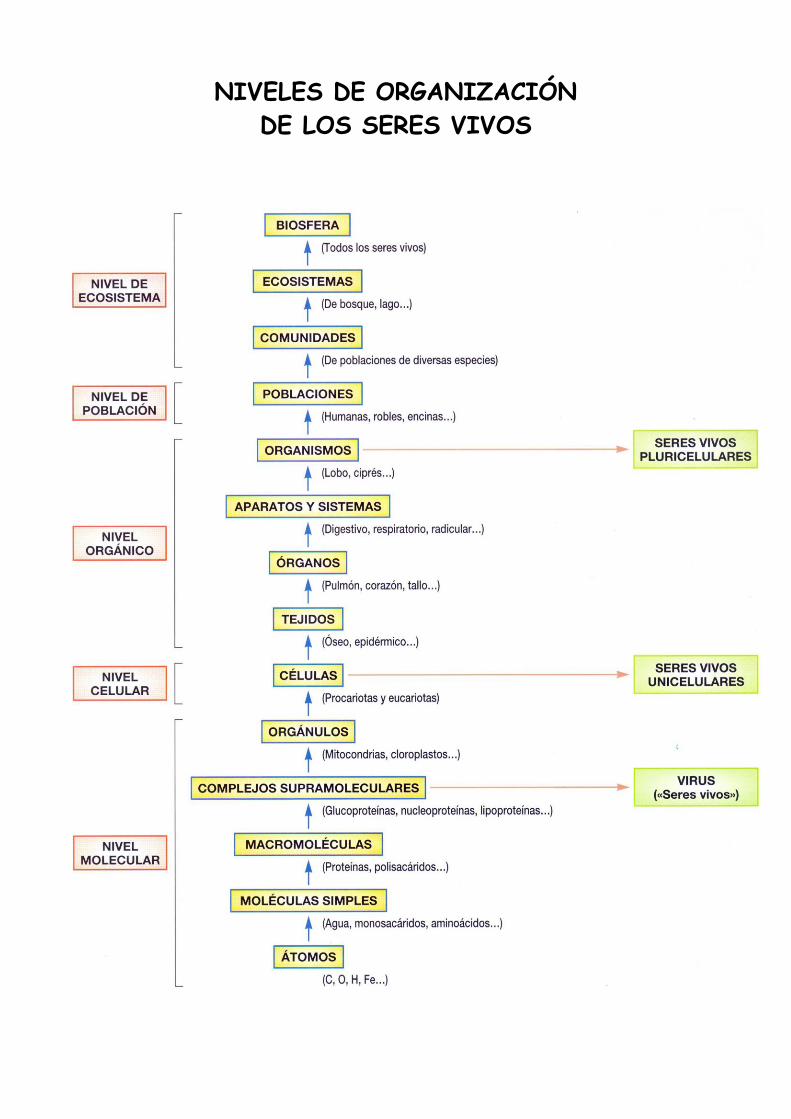
NIVELES DE ORGANIZACIÓN
DE LOS SERES VIVOS

La materia viva se encuentra estructurada en diferentes niveles de organización o grados de
complejidad que le proporcionan unas propiedades que no se dan en los niveles inferiores.
Dichos niveles son los siguientes:
Nivel Subatómico: Es aquel integrado por las partículas más pequeñas de la materia:
protones, neutrones y electrones.
Nivel atómico: Está representado por los átomos: parte más pequeña de un elemento
químico que puede intervenir en una reacción. Ejemplos: Fe, H, O, Cl,...
Nivel Molecular: Es el representado por moléculas: unidades materiales formadas por la
agrupación de 2 o más átomos mediante enlaces químicos. Dichas moléculas reciben el
nombre de moléculas orgánicas si están formadas por átomos de C unidos covalentemente.
Las moléculas orgánicas pueden ser, a su vez, de dos tipos:
o Biomoléculas o Principios Inmediatos: A este grupo pertenecen todas aquellas moléculas
orgánicas de C presentes en los seres vivos. Ej.: glucosa (C6H12O6)
o No Biomoléculas: Son aquellas moléculas orgánicas formadas por C y sintetizadas
artificialmente, no presentes en la materia viva. Ejemplo: los plásticos.
Dentro del nivel molecular, existen varios grados de complejidad o subniveles:
a) Subnivel Macromolecular: Está representado por las macromoléculas: polímeros de
alto Pm resultantes de la unión de muchas moléculas orgánicas (monómeros). Ejemplos
de polímeros: las proteínas, los ácidos nucleicos o el almidón. Ejemplos de monómeros:
los aminoácidos, los nucleótidos o la glucosa.
b) Subnivel de Complejo Supramolecular: Está representado por los complejos
supramoleculares: estructuras resultantes de la unión de varias macromoléculas. Un
ejemplo es el caso de las glucoproteínas, formadas por glúcidos y proteínas.
c) Subnivel de Orgánulo Celular: Los orgánulos celulares son pequeñas estructuras de
las células animales y vegetales con función propia, resultantes de la asociación de
complejos supramoleculares. Ejemplos: lisosomas, mitocondrias, centriolos, complejo
de Golgi, etc.
Nivel Celular: Es el representado por las células. Según la Teoría Celular de Virchov, una
célula es la “unidad vital, morfológica, fisiológica y genética de todos los seres vivos”. El
significado de dicha expresión es el siguiente:
- Unidad vital: la célula es el ser vivo menor y más sencillo.
- Unidad morfológica, anatómica o estructural: todo ser vivo está compuesto por una o
varias células.
- Unidad fisiológica o funcional: cada célula posee su propia actividad vital.
- Unidad genética o de origen: toda célula procede de otra preexistente.

Se distinguen dos tipos de células:
a) Células Procariotas: son las que carecen de envoltura nuclear, de modo que la
información genética está dispersa en el citoplasma. A este grupo pertenecen las
bacterias.
b) Células Eucariotas: son aquellas cuya información genética está rodeada por una
envoltura membranosa. A este grupo pertenecen las células animales y vegetales.
Las células pueden desarrollarse como organismos independientes o agruparse, de modo que
podemos hablar de:
1- Organismos Unicelulares, formados por una sola célula capaz de realizar todas las
funciones básicas de cualquier ser vivo (funciones de nutrición, relación y
reproducción).
2- Colonias: Son agrupaciones de células donde cada una realiza individualmente los tres
tipos de funciones.
3- Organismos pluricelulares, correspondientes al siguiente nivel de organización.
Nivel Pluricelular: Es aquel formado por más de una célula y a nivel del cual hay una
diferenciación o especialización funcional. Dentro de él existen varios subniveles o grados
de complejidad:
o Subnivel de Tejido. En este nivel, el tejido es definido como el conjunto de células
especializadas muy parecidas, que realizan la misma función y tienen el mismo origen. Si
el organismo pluricelular tiene u sólo tipo de células, entonces se dice que tiene
estructura de talo (propio de algas y hongos pluricelulares).
o Subnivel de Órgano. En él, el órgano es considerado como la unidad estructural y
funcional de los seres vivos superiores, formadas por varios tejidos diferentes que
realizan un ACTO concreto. Ejemplo: El corazón, formado por los tejidos muscular,
epitelial y nervioso.
o Subnivel de Sistema. Un sistema es considerado como un conjunto de órganos
parecidos, formados por los mismos tejidos, pero que realizan actos que pueden ser
independientes. Por ejemplo, dentro del sistema muscular existen músculos que mueven
la cabeza, otros que mueven los brazos, otros que actúan a nivel de las piernas, etc.
Nivel de Población, que se define como el conjunto de individuos de la misma especie, que
vive en un área y en un momento determinado.
Nivel de Ecosistema, que abarca la comunidad o biocenosis (conjunto de poblaciones de
distintos seres vivos que viven interrelacionados) y el biotopo (lugar y condiciones
fisicoquímicas bajo los que se desarrolla esa comunidad).
ECOSISTEMA = BIOCENOSIS + BIOTOPO
El conjunto de ecosistemas de toda la Tierra (Biosfera) es considerado como el nivel más
complejo de organización de los seres vivos.

CÉLULA. CONCEPTO, FORMA Y TAMAÑO
La célula es la estructura más simple conocida, capaz de realizar por sí misma las tres
funciones básicas de los seres vivos (nutrición, relación y reproducción), y constituida por
tres elementos básicos: la membrana plasmática, el citoplasma y el material genético (ADN).
De estas tres funciones vitales, la más importante es la reproducción, ya que es la que
caracteriza a un ser vivo, que puede ser definido como aquel capaz de engendrar a otros
seres vivos con esa misma capacidad.
Aunque los virus también tienen capacidad para reproducirse, sin embargo sólo lo pueden
hacer si disponen de una célula viva a la que infectar y por lo tanto, son incapaces de
sobrevivir por sí solos durante mucho tiempo. Así, podemos decir que, debido a su estrecha
dependencia de las células, los virus deberían ser considerados como materia viva, más que
como seres vivos. O dicho de otra forma: los virus están en la frontera entre los seres vivos
y la materia inerte. Los virus son una forma de vida acelular ya que no tienen estructura
celular, y por lo tanto, no pueden ser incluidos dentro del nivel celular, sino que deben ser
incluidos dentro del nivel anterior: el nivel macromolecular.
FORMA DE LAS CÉLULAS
Las células presentan gran variedad de formas e incluso, algunas carecen de forma fija. Las
células con formas definidas pueden ser redondeadas, elípticas, fusiformes, estrelladas,
prismáticas, aplanadas, etc.
Las células libres (amebas y leucocitos), con membrana plasmática fácilmente deformable y
sin membrana de secreción rígida, son capaces de variar su forma mediante la emisión de
pseudópodos (prolongaciones citoplasmáticas) que les permiten desplazarse y fagocitar
partículas. Sin embargo, otras células libres incapaces de emitir pseudópodos (ciliados,
eritrocitos, linfocitos) adoptan formas globosas, debido a la cohesión entre las moléculas de
agua.
Las células sin pared celular rígida y que forman parte de ciertos tejidos, tienen una forma
que depende de los fenómenos osmóticos y, en gran parte, de las tensiones que en ella generan
las uniones con las células contiguas. Se ha observado que si se separan las células de un tejido
rompiendo las conexiones que las unen y se colocan en un medio de cultivo, dichas células
tienden a adquirir la forma esférica.
Por su parte, las células con pared celular rígida (bacterias con pared de mureína, células
vegetales con pared de celulosa, osteocitos), aunque también están sometidas a fenómenos
osmóticos, tienen una forma muy estable.
En cualquier caso, la forma que adopte una célula está estrechamente relacionada con la
función que realiza. Así por ejemplo, las célula musculares son alargadas y fusiformes para
poderse contraer y relajar más fácilmente; las células del tejido nervioso tienen un aspecto
más o menos estrellado y con numerosas prolongaciones para poder captar y transmitir los
estímulos más fácilmente; los enterocitos tienen la membrana plasmática libre recubierta con
numerosos pliegues para aumentar la superficie de absorción de nutrientes; etc.
Por lo tanto, podemos decir que, aunque la forma de las células está claramente relacionada
con la función que realizan, ésta puede variar en función de la ausencia de pared celular rígida,
de las tensiones de uniones a células vecinas, de la viscosidad del citosol, de fenómenos
osmóticos e incluso, del tipo de citoesqueleto interno.

TAMAÑO DE LAS CÉLULAS
Al igual que con la forma, el tamaño celular
también es extremadamente variable. Así
por ejemplo, las bacterias miden entre 1 y 5
μ de longitud; la mayoría de células humanas
miden entre 5 y 20 μ; los espermatozoides
humanos miden unos 53 μ de longitud; el
oocito humano, unas 150 μ; los granos de
polen de ciertas plantas, entre 200 y 300 μ;
e incluso, las neuronas de los cetáceos
pueden llegar a alcanzar varios metros de
longitud a través de sus prolongaciones
axonales.
A pesar de esta diversidad, el tamaño celular
está controlado por dos factores básicos: la
capacidad de captación de nutrientes del
medio que rodea a la célula y la capacidad
funcional del núcleo.
UNIDADES DE MEDIDA
Aunque algunas células pueden ser
observadas a simple vista (huevos de las
aves), la mayoría sólo son observables al
microscopio y por tanto, debido a sus ínfimos
tamaños, sus dimensiones tienen que ser
dadas en unidades inferiores al milímetro.
Entre ellas, destacan las siguientes:
1 micrómetro (o micra)= 1 μm = 0,001 mm (milésima de milímetro)
1 nanómetro = 1 nm = 0,000 001 mm (millonésima de milímetro)
1 ángstrom = 1 Å = 0,1 nm (diez millonésimas de milímetro)
Cuando una célula tridimensional, por ejemplo, esférica, aumenta de tamaño, su volumen aumenta
proporcionalmente al cubo del radio (V = 4/3·Лr3), mientras que su superficie sólo aumenta en función del
cuadrado del radio (S = 4Лr2). Es decir, aumenta mucho más su volumen que su superficie y por lo tanto, la
relación superficie/volumen disminuye, lo que supone un gran inconveniente para la supervivencia de la célula,
ya que la entrada de todos los nutrientes necesarios para la célula está en función de su superficie y, si ésta
es insuficiente, la célula terminará por morir. Por este motivo, muy pocas células maduras son esféricas, sino
que son aplanadas, prismáticas o irregulares, con el fin de mantener la relación superficie/volumen
aproximadamente constante.
Por otra parte, el aumento del volumen celular no va acompañado de un aumento del volumen nuclear, ni de su
dotación cromosómica, sino que dentro de una misma estirpe celular, la forma globosa y la relación S/V
grande son típicas de células jóvenes. Así por ejemplo, cuanto más grandes son los oocitos (de formas
globosas), menor es la relación S/V y tanto más próximos estarán a su madurez, fecundación e inicio de su
división, o bien a su degradación. Así mismo, dentro de una misma estirpe celular, cuanto menor sea la
relación volumen nuclear/volumen citoplasmático, más próxima estará a dividirse, si tiene capacidad para
ello, o a morir, si no la tiene. En este caso, también hay que tener en cuenta el grado de empaquetamiento
de la cromatina. Así, una cromatina extendida que facilita la transcripción del ADN, se relaciona con una
célula en plena actividad metabólica, mientras que una cromatina muy empaquetada indica el inicio de división
celular o bien su muerte.
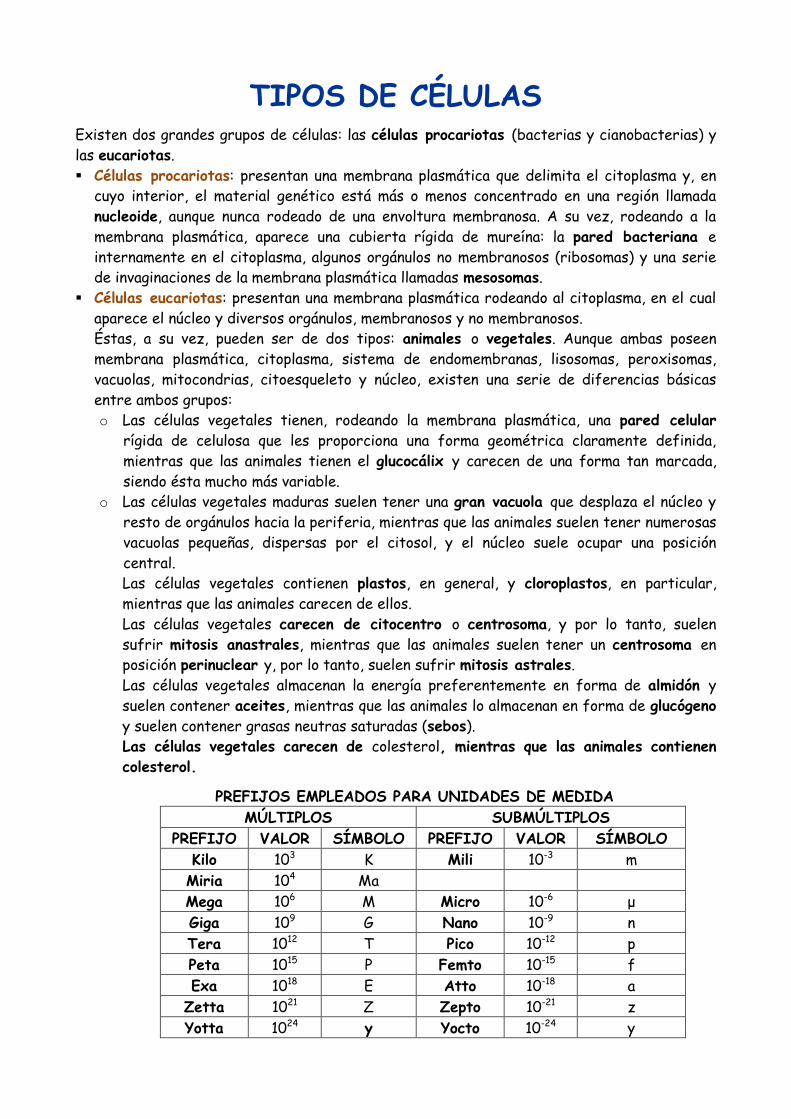
TIPOS DE CÉLULAS
Existen dos grandes grupos de células: las células procariotas (bacterias y cianobacterias) y
las eucariotas.
Células procariotas: presentan una membrana plasmática que delimita el citoplasma y, en
cuyo interior, el material genético está más o menos concentrado en una región llamada
nucleoide, aunque nunca rodeado de una envoltura membranosa. A su vez, rodeando a la
membrana plasmática, aparece una cubierta rígida de mureína: la pared bacteriana e
internamente en el citoplasma, algunos orgánulos no membranosos (ribosomas) y una serie
de invaginaciones de la membrana plasmática llamadas mesosomas.
Células eucariotas: presentan una membrana plasmática rodeando al citoplasma, en el cual
aparece el núcleo y diversos orgánulos, membranosos y no membranosos.
Éstas, a su vez, pueden ser de dos tipos: animales o vegetales. Aunque ambas poseen
membrana plasmática, citoplasma, sistema de endomembranas, lisosomas, peroxisomas,
vacuolas, mitocondrias, citoesqueleto y núcleo, existen una serie de diferencias básicas
entre ambos grupos:
o Las células vegetales tienen, rodeando la membrana plasmática, una pared celular
rígida de celulosa que les proporciona una forma geométrica claramente definida,
mientras que las animales tienen el glucocálix y carecen de una forma tan marcada,
siendo ésta mucho más variable.
o Las células vegetales maduras suelen tener una gran vacuola que desplaza el núcleo y
resto de orgánulos hacia la periferia, mientras que las animales suelen tener numerosas
vacuolas pequeñas, dispersas por el citosol, y el núcleo suele ocupar una posición
central.
Las células vegetales contienen plastos, en general, y cloroplastos, en particular,
mientras que las animales carecen de ellos.
Las células vegetales carecen de citocentro o centrosoma, y por lo tanto, suelen
sufrir mitosis anastrales, mientras que las animales suelen tener un centrosoma en
posición perinuclear y, por lo tanto, suelen sufrir mitosis astrales.
Las células vegetales almacenan la energía preferentemente en forma de almidón y
suelen contener aceites, mientras que las animales lo almacenan en forma de glucógeno
y suelen contener grasas neutras saturadas (sebos).
Las células vegetales carecen de colesterol, mientras que las animales contienen
colesterol.
PREFIJOS EMPLEADOS PARA UNIDADES DE MEDIDA
MÚLTIPLOS SUBMÚLTIPLOS
PREFIJO VALOR SÍMBOLO PREFIJO VALOR SÍMBOLO
Kilo 103 K Mili 10-3 m
Miria 104 Ma
Mega 106 M Micro 10-6
μ
Giga 109 G Nano 10-9
n
Tera 1012 T Pico 10-12
p
Peta 1015 P Femto 10-15
f
Exa 1018 E Atto 10-18
a
Zetta 1021 Z Zepto 10-21
z
Yotta 1024 y Yocto 10-24
y
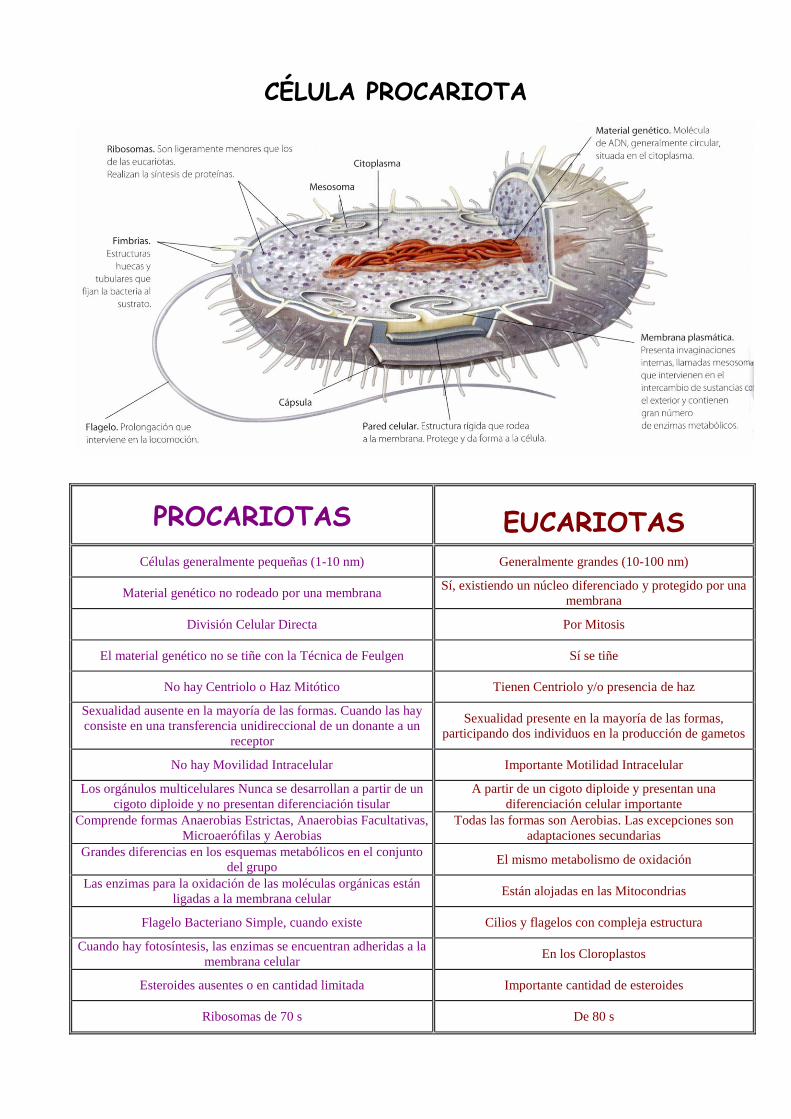
CÉLULA PROCARIOTA
PROCARIOTAS EUCARIOTAS Células generalmente pequeñas (1-10 nm) Generalmente grandes (10-100 nm)
Material genético no rodeado por una membrana Sí, existiendo un núcleo diferenciado y protegido por una
membrana
División Celular Directa Por Mitosis
El material genético no se tiñe con la Técnica de Feulgen Sí se tiñe
No hay Centriolo o Haz Mitótico Tienen Centriolo y/o presencia de haz
Sexualidad ausente en la mayoría de las formas. Cuando las hay
consiste en una transferencia unidireccional de un donante a un
receptor
Sexualidad presente en la mayoría de las formas,
participando dos individuos en la producción de gametos
No hay Movilidad Intracelular Importante Motilidad Intracelular
Los orgánulos multicelulares Nunca se desarrollan a partir de un
cigoto diploide y no presentan diferenciación tisular
A partir de un cigoto diploide y presentan una
diferenciación celular importante
Comprende formas Anaerobias Estrictas, Anaerobias Facultativas,
Microaerófilas y Aerobias
Todas las formas son Aerobias. Las excepciones son
adaptaciones secundarias
Grandes diferencias en los esquemas metabólicos en el conjunto
del grupo El mismo metabolismo de oxidación
Las enzimas para la oxidación de las moléculas orgánicas están
ligadas a la membrana celular Están alojadas en las Mitocondrias
Flagelo Bacteriano Simple, cuando existe Cilios y flagelos con compleja estructura
Cuando hay fotosíntesis, las enzimas se encuentran adheridas a la
membrana celular En los Cloroplastos
Esteroides ausentes o en cantidad limitada Importante cantidad de esteroides
Ribosomas de 70 s De 80 s
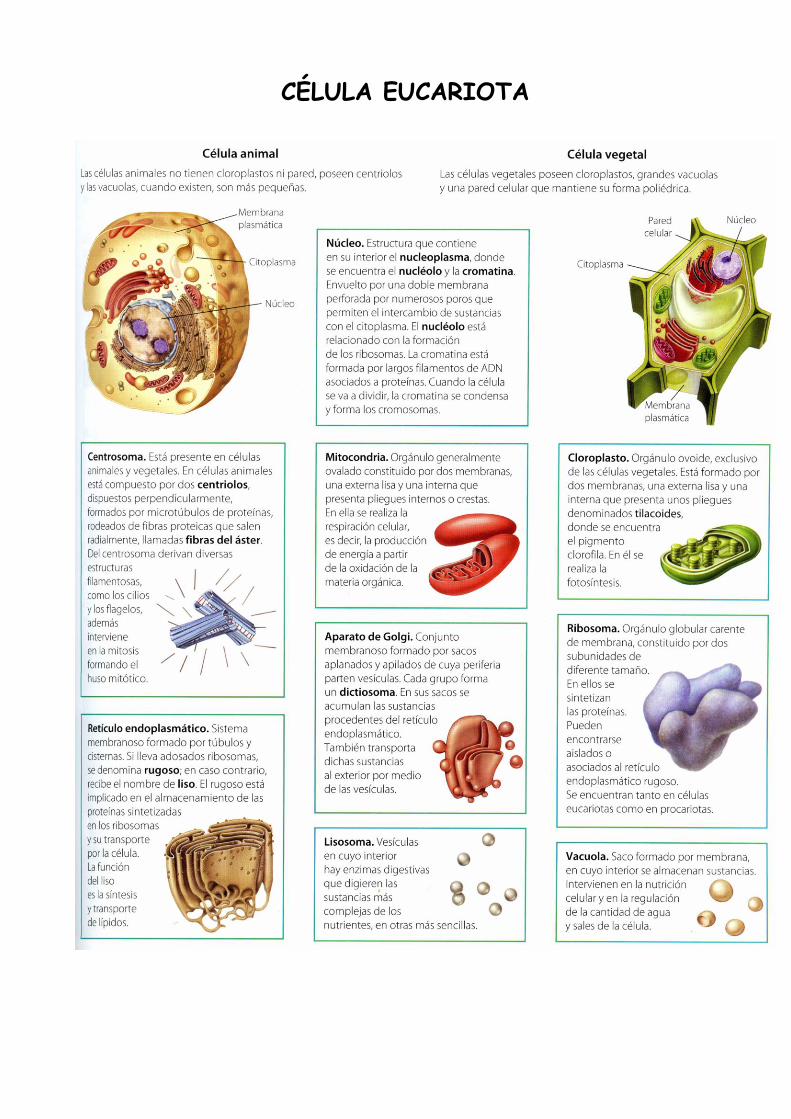
CÉLULA EUCARIOTA
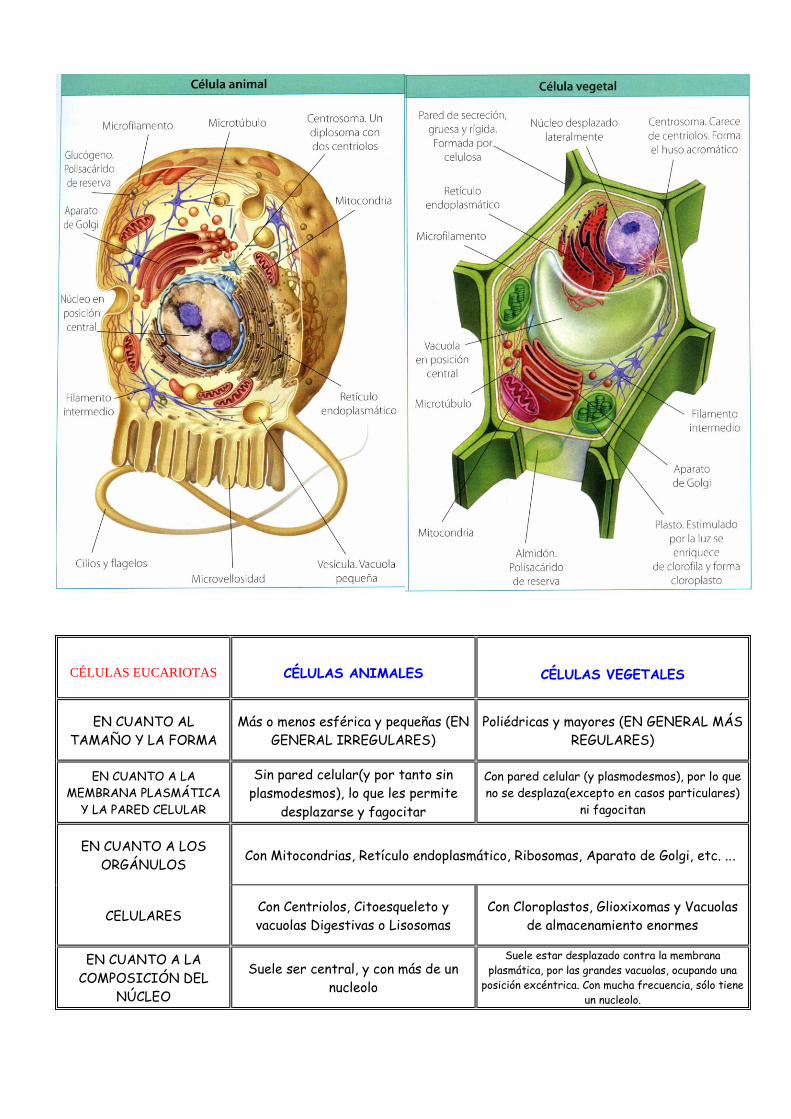
CÉLULAS EUCARIOTAS
CÉLULAS ANIMALES CÉLULAS VEGETALES
EN CUANTO AL
TAMAÑO Y LA FORMA
Más o menos esférica y pequeñas (EN
GENERAL IRREGULARES)
Poliédricas y mayores (EN GENERAL MÁS
REGULARES)
EN CUANTO A LA
MEMBRANA PLASMÁTICA
Y LA PARED CELULAR
Sin pared celular(y por tanto sin
plasmodesmos), lo que les permite
desplazarse y fagocitar
Con pared celular (y plasmodesmos), por lo que
no se desplaza(excepto en casos particulares)
ni fagocitan
EN CUANTO A LOS
ORGÁNULOS Con Mitocondrias, Retículo endoplasmático, Ribosomas, Aparato de Golgi, etc. ...
CELULARES Con Centriolos, Citoesqueleto y
vacuolas Digestivas o Lisosomas
Con Cloroplastos, Glioxixomas y Vacuolas
de almacenamiento enormes
EN CUANTO A LA
COMPOSICIÓN DEL
NÚCLEO
Suele ser central, y con más de un
nucleolo
Suele estar desplazado contra la membrana
plasmática, por las grandes vacuolas, ocupando una
posición excéntrica. Con mucha frecuencia, sólo tiene
un nucleolo.

DIFERENCIAS ENTRE CÉLULA PROCARIOTA Y CÉLULA EUCARIOTA
● La célula procariota no tiene núcleo protector del material genético. La célula eucariota sí presenta
núcleo limitado por una estructura membranosa.
● El citoplasma de la célula eucariota se encuentra compartimentado, presentando orgánulos, mientras
que en la procariota no aparece esta compartimentación.
● La célula procariota está protegida por una pared bacteriana distinta a la que envuelve a las células
vegetales.
● Las células procariotas son organismos más primitivos que las células eucariotas.
● El ADN de células procariotas es circular,
mientras que el ADN de eucariotas es lineal.
● Cuando presentan flagelos, la estructura es
diferente en procariotas y eucariotas.
● La membrana plasmática de procariotas contiene
más cantidad de proteínas que la membrana de
las eucariotas.
● La célula procariota tiene invaginaciones en su
membrana, denominadas mesosomas.
DIFERENCIAS ENTRE CÉLULA VEGETAL Y CÉLULA ANIMAL
Las plantas están formadas por células, igual que los animales. Sin embargo, aunque la mayor parte de
estructuras celulares que presentan funcionan igual, tienen otras estructuras distintas a las de las
células animales:
● La célula vegetal suele ser más grande que la animal y presenta una pared exterior, rígida, formada
por celulosa. La pared celular da a la célula vegetal una forma constante y con aspecto poliédrico.
● Además, la célula vegetal contiene orgánulos fotosintéticos, de color verde por su contenido en
clorofila. Estos orgánulos se llaman cloroplastos. Las células con cloroplastos son autótrofas. Sin
embargo, las células animales son heterótrofas.
● Las células vegetales suelen contener una bolsa de almacén de sustancias, principalmente agua. A
este orgánulo se le llama vacuola, que a veces es tan grande que ocupa casi todo el interior celular.
● Las células vegetales no presentan centriolos, aunque tienen citoesqueleto y forman huso acromático
en su división.
● Cuando la célula se divide en dos, por mitosis, la división del citoplasma se realiza por la formación de
una pared que la separa en dos partes, se conoce con el nombre de fragmoplasto. El resultado, igual
que en la célula animal, es la formación de dos células hijas, idénticas entre sí e idénticas a la célula
madre.
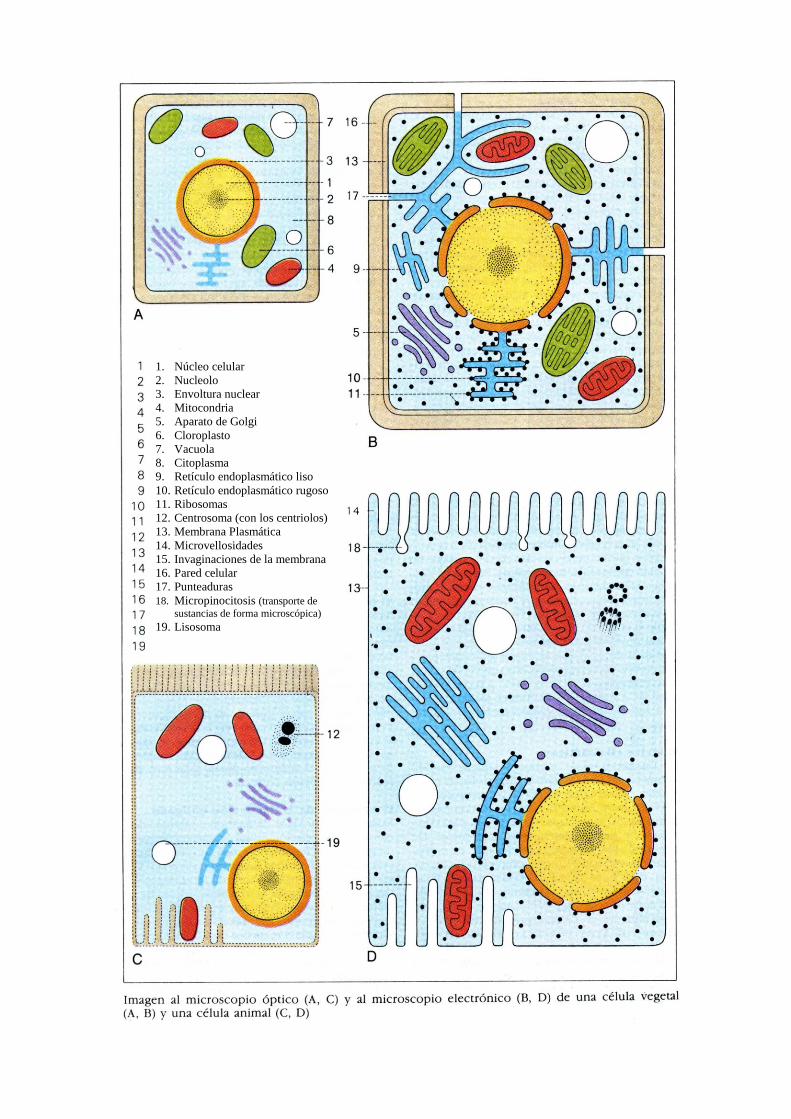
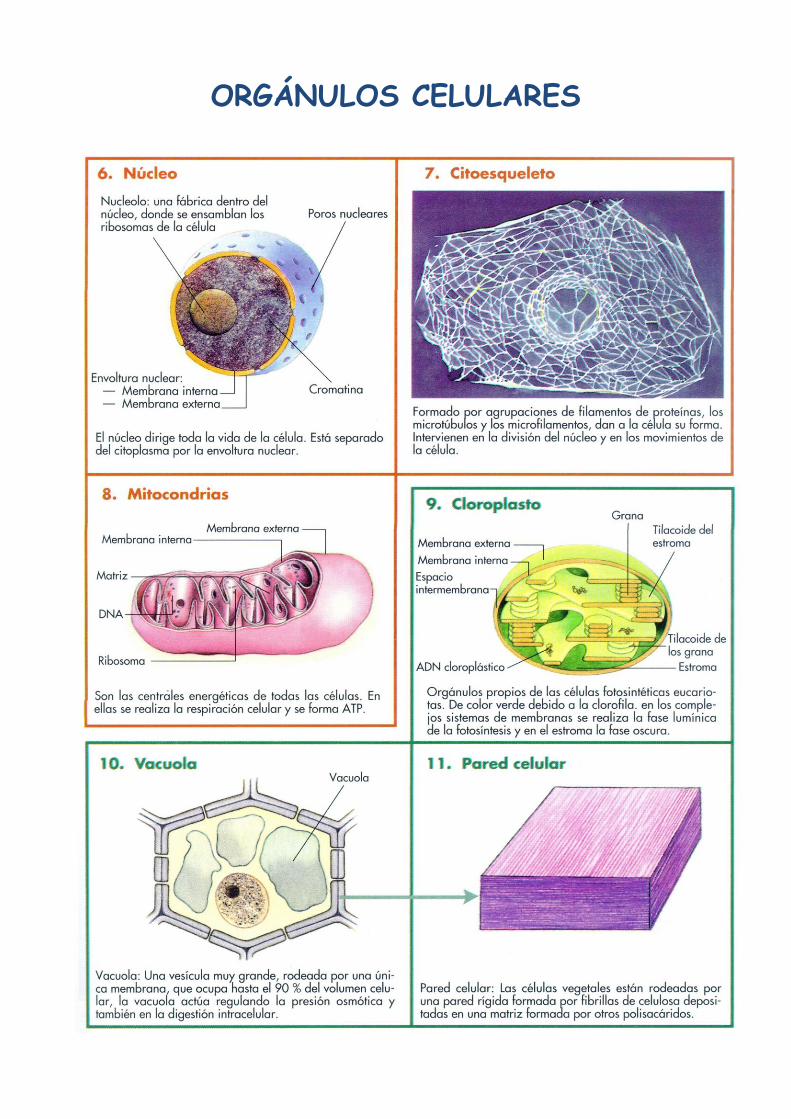
1. Núcleo celular
2. Nucleolo
3. Envoltura nuclear
4. Mitocondria
5. Aparato de Golgi
6. Cloroplasto
7. Vacuola
8. Citoplasma
9. Retículo endoplasmático liso
10. Retículo endoplasmático rugoso
11. Ribosomas
12. Centrosoma (con los centriolos)
13. Membrana Plasmática
14. Microvellosidades
15. Invaginaciones de la membrana
16. Pared celular
17. Punteaduras
18. Micropinocitosis (transporte de
sustancias de forma microscópica)
19. Lisosoma



PARED CELULAR DEFINICIÓN Es una capa rígida que rodea y protege a la membrana plasmática de las células vegetales, algas y hongos, consiguiendo
así un sostén indispensable.
COMPOSICIÓN Pared Primaria: Fibras de Celulosa orientadas en muchas y diferentes direcciones, formando una red
relativamente, laxa, y Hemicelulosa en el caso de los vegetales y algas y Quitina en el caso de los hongos.
Pared Secundaria: Fibras de Celulosa paralelas, dispuestas en capas alternadas que le confiere menor flexibilidad
y elasticidad; Lignina que le otorga gran resistencia y Pectinas.
Lámina Media (puede aparecer entre paredes): Pectatos de Ca y Mg, con propiedades cementantes.
FUNCIÓN Otorga Rigidez, Soporte, Resistencia y Protección a la célula.
Es capaz de extenderse para que la célula crezca.
Es permeable a todas las moléculas.
DIFERENCIACIONES PUNTEADURAS: Adelgazamientos de la pared celular que generalmente se corresponden con otras
complementarias y al mismo nivel de la célula adyacente.
PLASMODESMOS: Finos canales que atraviesan totalmente las paredes celulares, estableciendo continuidad
entre los citoplasmas de células contiguas.

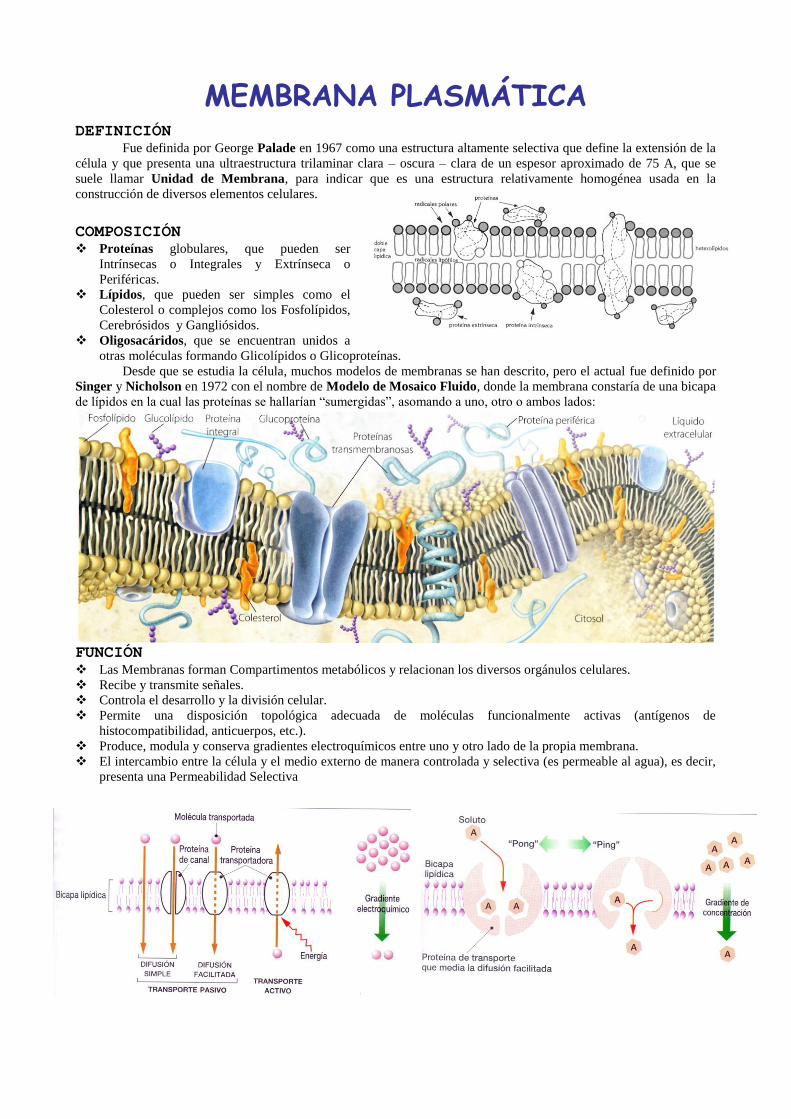
MEMBRANA PLASMÁTICA
DEFINICIÓN Fue definida por George Palade en 1967 como una estructura altamente selectiva que define la extensión de la
célula y que presenta una ultraestructura trilaminar clara – oscura – clara de un espesor aproximado de 75 A, que se
suele llamar Unidad de Membrana, para indicar que es una estructura relativamente homogénea usada en la
construcción de diversos elementos celulares.
COMPOSICIÓN Proteínas globulares, que pueden ser
Intrínsecas o Integrales y Extrínseca o
Periféricas.
Lípidos, que pueden ser simples como el
Colesterol o complejos como los Fosfolípidos,
Cerebrósidos y Gangliósidos.
Oligosacáridos, que se encuentran unidos a
otras moléculas formando Glicolípidos o Glicoproteínas.
Desde que se estudia la célula, muchos modelos de membranas se han descrito, pero el actual fue definido por
Singer y Nicholson en 1972 con el nombre de Modelo de Mosaico Fluido, donde la membrana constaría de una bicapa
de lípidos en la cual las proteínas se hallarían “sumergidas”, asomando a uno, otro o ambos lados:
FUNCIÓN
Las Membranas forman Compartimentos metabólicos y relacionan los diversos orgánulos celulares.
Recibe y transmite señales.
Controla el desarrollo y la división celular.
Permite una disposición topológica adecuada de moléculas funcionalmente activas (antígenos de
histocompatibilidad, anticuerpos, etc.).
Produce, modula y conserva gradientes electroquímicos entre uno y otro lado de la propia membrana.
El intercambio entre la célula y el medio externo de manera controlada y selectiva (es permeable al agua), es decir,
presenta una Permeabilidad Selectiva


TRANSPORTE Y MEMBRANA PLASMÁTICA
Moléculas de pequeño tamaño:
Transporte Pasivo (realizado a favor de gradiente y sin consumo de energía
metabólica) por Difusión Simple o Difusión Facilitada.
Transporte Activo, contra gradiente y con consumo de energía metabólica.
Cotransporte, en el que a la vez que se introduce una molécula, otra es sacada.
Moléculas de gran tamaño:
Endocitosis (cuando se introduce materia en el citoplasma), que puede ser Fagocitosis
si se trata de microorganismos o sustancias sólidas y Pinoctiosis si se trata de
macromoléculas o sustancias líquidas.
Exocitosis cuando se transporta material desde el interior de la célula al medio
extracelular.
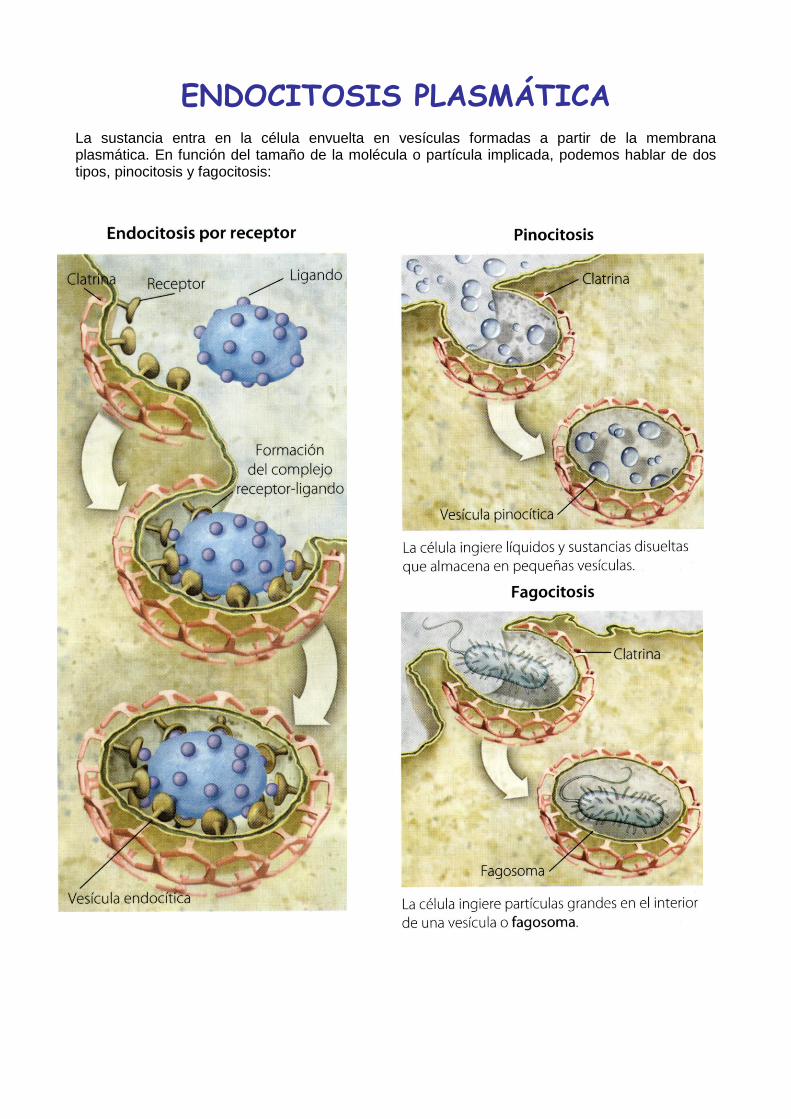
ENDOCITOSIS PLASMÁTICA
La sustancia entra en la célula envuelta en vesículas formadas a partir de la membrana plasmática. En función del tamaño de la molécula o partícula implicada, podemos hablar de dos tipos, pinocitosis y fagocitosis:

EXOCITOSIS PLASMÁTICA
La exocitosis es la fusión de vesículas producidas principalmente por el aparato de Golgi con
la membrana plasmática. Las vesículas se forman en la cara trans del aparato de Golgi y viajan
hasta la membrana plasmática con quien se fusionan.
Hay dos tipos de exocitosis: constitutiva y regulada.
La exocitosis constitutiva se produce en todas las células y se encarga de liberar
moléculas que van a formar parte de la matriz extracelular o bien sirven para regenerar la
propia membrana celular. Es un proceso constante de producción, desplazamiento y fusión,
con diferente intensidad de tráfico según el estado fisiológico de la célula.
La exocitosis regulada se produce sólo en aquellas células especializadas en la secreción,
como por ejemplo las productoras de hormonas, las neuronas, las células del epitelio
digestivo, las células glandulares y otras. En este tipo de exocitosis se liberan moléculas
que realizan funciones para el organismo como la digestión o que afectan a la fisiología de
otras células próximas o localizadas en regiones alejadas en el organismo, a las cuales llegan
a través del sistema circulatorio, como es el caso de las hormonas. Las vesículas de
secreción regulada no se fusionan espontáneamente con la membrana plasmática sino que
necesitan una señal consistente en el aumento de la concentración de calcio. Además,
necesitan ATP y GTP.
Las vesículas de la secreción regulada se acumulan en el citoplasma y cuando reciben la
señal para su liberación se dirigen hacia regiones concretas de la membrana de la célula,
luego es un proceso dirigido en el tiempo y en el espacio. Las células nerviosas representan
un ejemplo extremo. Una vez empaquetadas las vesículas en el soma neuronal tienen que ser
dirigidas hacia el terminal presináptico, que en algunas neuronas puede estar a centímetros
de distancia. Además de las neuronas existen otras células polarizadas, como es el caso de
las del epitelio digestivo, que poseen una parte apical y otra basal. Sería un desastre que
las células epiteliales intestinales fusionasen las vesículas y liberasen las enzimas digestivas
que contienen en la región de la membrana celular orientada hacia los tejidos internos y no
hacia la luz del tubo digestivo.
La direccionalidad del camino de estas vesículas está determinada por la orientación del
citoesqueleto, el cual, mediante la intervención de las proteínas motoras, las transporta
hasta su lugar de fusión apropiado.
Estos dos tipos de exocitosis, constitutiva y regulada, empaquetan moléculas diferentes, luego
la cara trans del complejo de Golgi debe arreglárselas para separar ambos tipos de cargas.
Parece ser que las moléculas que no tienen una señal específica serán empaquetadas en
vesículas de exocitosis constitutiva. Las proteínas del interior de estas vesículas son liberadas
en el medio extracelular mientras que las integrales formarán parte de la membrana
plasmática. En el caso de las vesículas de secreción regulada se forman inicialmente pequeñas
vesículas que una vez en el citosol se fusionan entre sí para formar otras más grandes que
permanecen en el interior celular hasta que llega una señal que permite la fusión con la
membrana plasmática.

MODO DE SELECCIÓN DE MOLÉCULAS PARA LAS
VESÍCULAS DE EXOCITOSIS REGULADA
Este mecanismo se basa en la formación de agregados moleculares formados por las moléculas
que serán liberadas y que tienen actividad fisiológica, así como por las enzimas que se
encargan de su procesamiento. Muchas de estas moléculas que se liberan por exocitosis
regulada son incorporadas a las vesículas en formas no activas, por ejemplo propéptidos, que
son procesadas a sus formas activas una vez que las vesículas se han formado. Estos
agregados están formados por moléculas que no han sido secuestradas por las vesículas
cubiertas de clatrina, que van a los endosomas, ni por las vesículas cubiertas por COP-I, que se
dirigen hacia el lado cis del aparato de Golgi.
La liberación de moléculas al exterior celular supone la fusión de la membrana de la vesícula
con la membrana plasmática, de la cual terminará por formar parte. Sin embargo, en base a las
imágenes obtenidas con el microscopio electrónico se ha propuesto otro modelo de exocitosis
denominado "besa y corre" (kiss-and-run). Aquí la vesícula no se fusiona completamente con la
membrana sino que lo hace de una manera incompleta formando un poro que comunica el
interior de la vesícula con el exterior celular por donde liberará su contenido. Posteriormente
se cierra el poro quedando la vesícula vacía en el citosol. Este tipo de exocitosis se ha
propuesto para las sinapsis y para las células cromafines.
No todas las vesículas que se fusionan con la membrana plasmática provienen del aparato de
Golgi. Los endosomas tempranos son orgánulos especializados en recibir vesículas formadas en
la membrana plasmática, proceso denominado endocitosis. Tras su fusión con el endosoma
parte del contenido vesicular es reciclado y llevado de vuelta a la membrana plasmática por
medio de vesículas que se forman en el propio endosoma. Otro ejemplo lo tenemos en los
terminales sinápticos del sistema nervioso. Estos sitios de exocitosis están muy alejados del
aparato de Golgi, localizado en el soma neuronal. La liberación de neurotransmisores en la
sinapsis no puede depender en exclusiva del empaquetado de éstos en el TGN, sería una
comunicación nerviosa muy ineficiente y demasiado lenta. En los terminales nerviosos existe
un reciclado de vesículas que permite una exocitosis permanente e independiente del TGN. En
el terminal presináptico se produce la exocitosis en la zona de liberación, mientras que en la
membrana plasmática lateral del propio terminal se producen vesículas por invaginación que se
volverán a llenar con neurotransmisores gracias a la existencia de transportadores
específicos en sus membranas. Estas vesículas rellenadas sufren un nuevo proceso de
exocitosis. Normalmente liberan neurotransmisores pequeños con vías de síntesis poco
complejas. De este modo existe un proceso constante de exocitosis y formación de vesículas
localizado en el propio terminal presináptico.
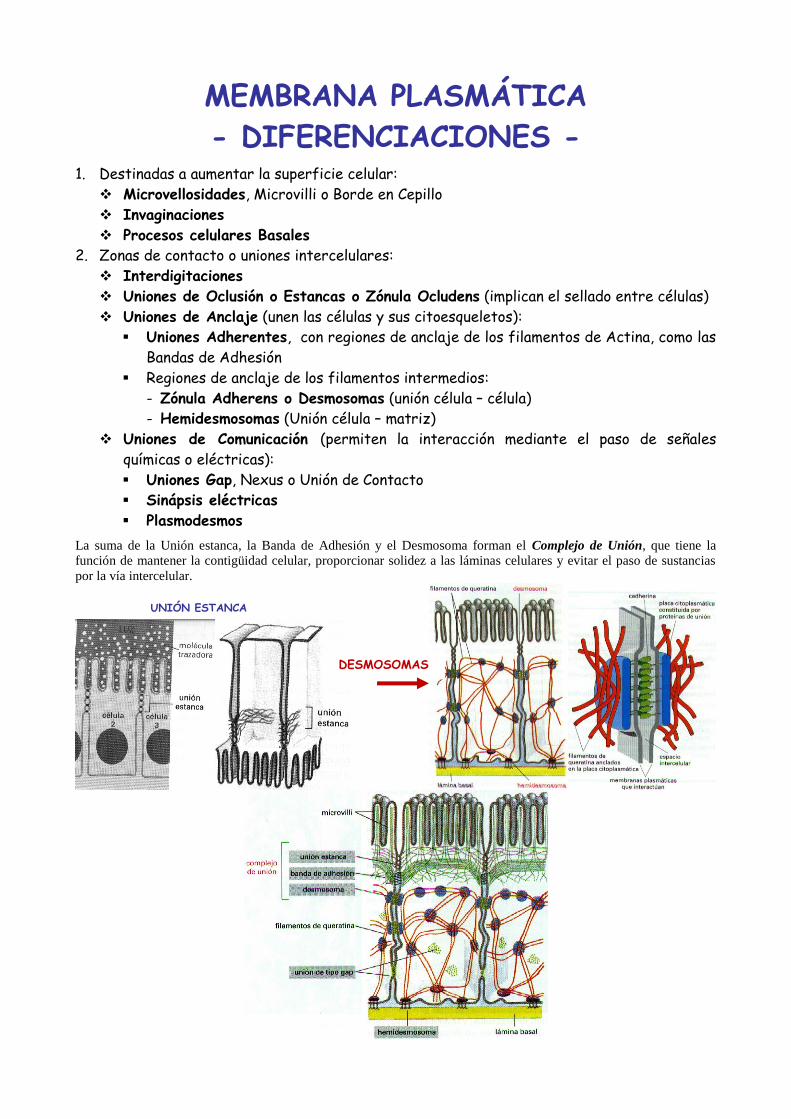
MEMBRANA PLASMÁTICA
- DIFERENCIACIONES -
1. Destinadas a aumentar la superficie celular:
Microvellosidades, Microvilli o Borde en Cepillo
Invaginaciones
Procesos celulares Basales
2. Zonas de contacto o uniones intercelulares:
Interdigitaciones
Uniones de Oclusión o Estancas o Zónula Ocludens (implican el sellado entre células)
Uniones de Anclaje (unen las células y sus citoesqueletos):
Uniones Adherentes, con regiones de anclaje de los filamentos de Actina, como las
Bandas de Adhesión
Regiones de anclaje de los filamentos intermedios:
- Zónula Adherens o Desmosomas (unión célula – célula)
- Hemidesmosomas (Unión célula – matriz)
Uniones de Comunicación (permiten la interacción mediante el paso de señales
químicas o eléctricas):
Uniones Gap, Nexus o Unión de Contacto
Sinápsis eléctricas
Plasmodesmos
La suma de la Unión estanca, la Banda de Adhesión y el Desmosoma forman el Complejo de Unión, que tiene la
función de mantener la contigüidad celular, proporcionar solidez a las láminas celulares y evitar el paso de sustancias
por la vía intercelular.
UNIÓN ESTANCA
DESMOSOMAS
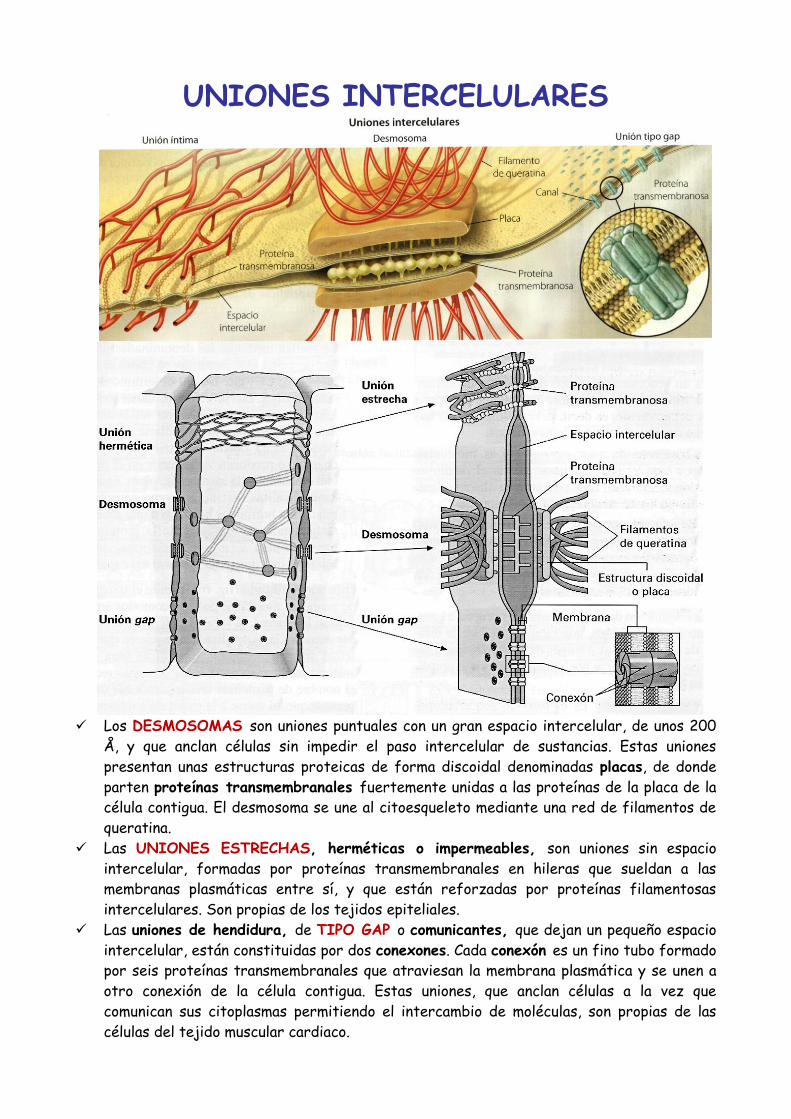
UNIONES INTERCELULARES
Los DESMOSOMAS son uniones puntuales con un gran espacio intercelular, de unos 200
Å, y que anclan células sin impedir el paso intercelular de sustancias. Estas uniones
presentan unas estructuras proteicas de forma discoidal denominadas placas, de donde
parten proteínas transmembranales fuertemente unidas a las proteínas de la placa de la
célula contigua. El desmosoma se une al citoesqueleto mediante una red de filamentos de
queratina.
Las UNIONES ESTRECHAS, herméticas o impermeables, son uniones sin espacio
intercelular, formadas por proteínas transmembranales en hileras que sueldan a las
membranas plasmáticas entre sí, y que están reforzadas por proteínas filamentosas
intercelulares. Son propias de los tejidos epiteliales.
Las uniones de hendidura, de TIPO GAP o comunicantes, que dejan un pequeño espacio
intercelular, están constituidas por dos conexones. Cada conexón es un fino tubo formado
por seis proteínas transmembranales que atraviesan la membrana plasmática y se unen a
otro conexión de la célula contigua. Estas uniones, que anclan células a la vez que
comunican sus citoplasmas permitiendo el intercambio de moléculas, son propias de las
células del tejido muscular cardiaco.
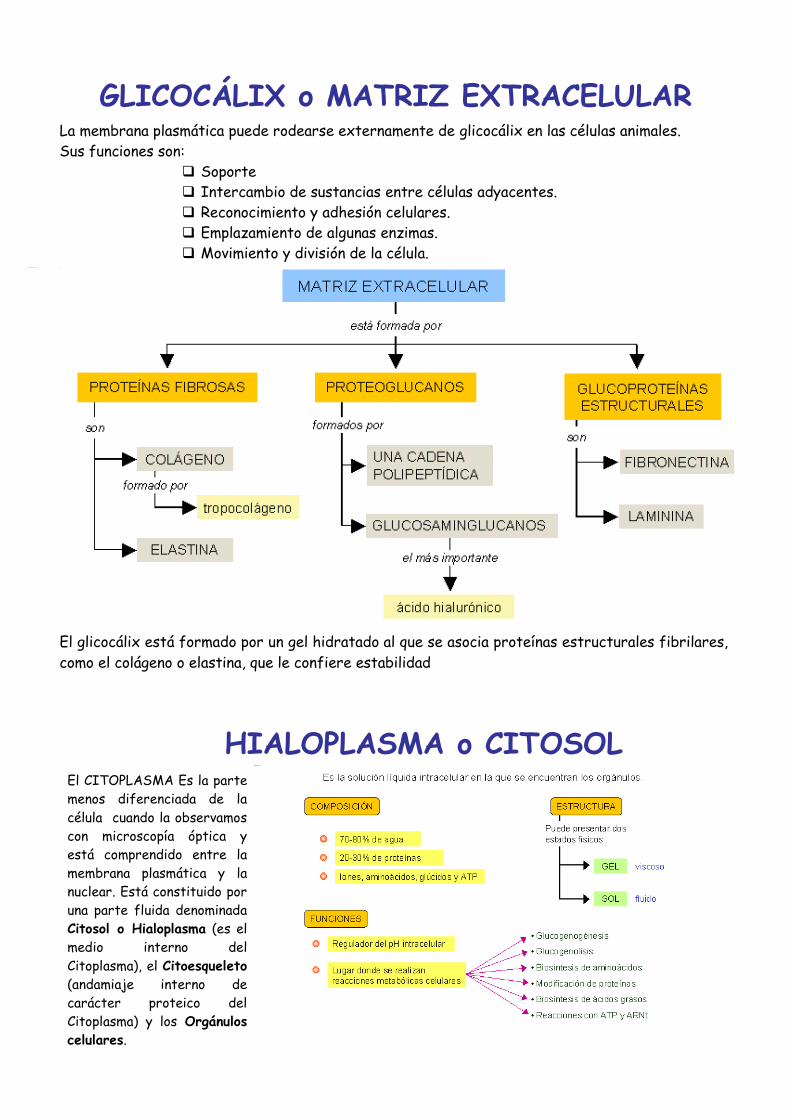
GLICOCÁLIX o MATRIZ EXTRACELULAR
La membrana plasmática puede rodearse externamente de glicocálix en las células animales.
Sus funciones son:
Soporte
Intercambio de sustancias entre células adyacentes.
Reconocimiento y adhesión celulares.
Emplazamiento de algunas enzimas.
Movimiento y división de la célula.
El glicocálix está formado por un gel hidratado al que se asocia proteínas estructurales fibrilares,
como el colágeno o elastina, que le confiere estabilidad
HIALOPLASMA o CITOSOL
El CITOPLASMA Es la parte
menos diferenciada de la
célula cuando la observamos
con microscopía óptica y
está comprendido entre la
membrana plasmática y la
nuclear. Está constituido por
una parte fluida denominada
Citosol o Hialoplasma (es el
medio interno del
Citoplasma), el Citoesqueleto
(andamiaje interno de
carácter proteico del
Citoplasma) y los Orgánulos
celulares.

CITOESQUELETO
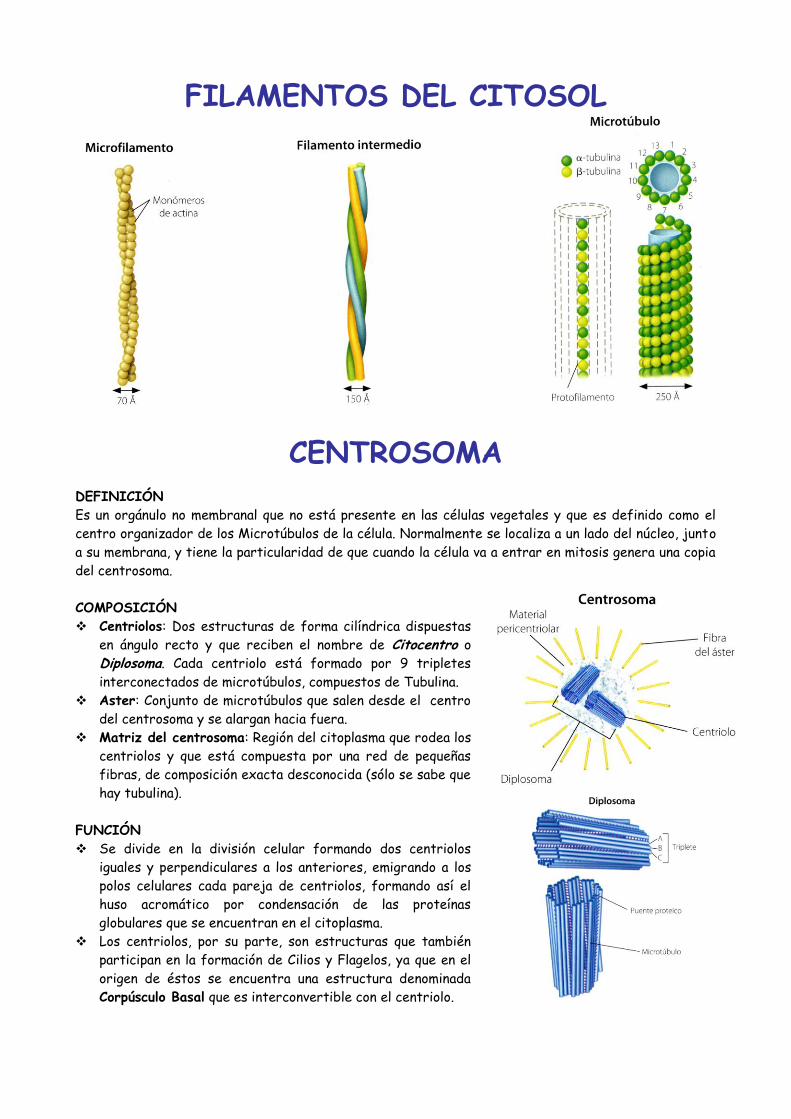
COMPOSICIÓN
Microfilamentos: De Actina.
Filamentos Intermedios: Varios
tipos de proteínas, como
Queratinas, De Desmina,
Gliales, etc.
Microtúbulos: De Tubulina,
pudiendo existir Lábiles y
Estables.
Proteínas accesorias que
participan en el ensamblaje de
aquellos y/o en su movimiento.
FUNCIONES GENERALES
Mantenimiento de la forma celular
Del posicionamiento y desplazamiento intracelular de los orgánulos.
Del movimiento celular.
División celular.
ESQUELETO MECÁNICO: Dando rigidez a las microvellosidades
de células epiteliales y actuando como Receptores de vibraciones en
los estereocilios de células ciliadas del oído interno.
FUERZA MECÁNICA DE CARÁCTER CONTRÁCTIL: Actúan
en la contractibilidad muscular en asociación con fibras de Miosina;
Junto a fibras de Miosina participan en el proceso de la División
Celular formando el anillo contráctil que conduce a la Citocinesis;
Generando cierta invaginación membranal en los Desmosomas;
Participando en la Locomoción ameboide y en los fenómenos de
tipo ameboide en general.
PROPORCIONAN ESTABILIDAD MECÁNICA
COLABORAN en la formación del Retículo del Citoesqueleto,
participando en sus funciones.
TRANSPORTE INTRACELULAR: Controlando la circulación de
sustancias en el citoplasma.
INVOLUCRACIÓN EN LA MORFOGÉNESIS: Colaboran en la
adquisición de la adecuada forma celular, como necesidad para el
desarrollo de la función asignada.
ADQUISICIÓN DE POLARIDAD CELULAR: Cuando las
funciones de la célula guardan relación con las distintas superficies
de la misma.
INTERVENCIÓN EN LA MOTILIDAD
COMPONENTES DE CENTRIOLOS, FLAGELOS Y CILIOS
FILAMENTOS DEL CITOSOL
CENTROSOMA
DEFINICIÓN
Es un orgánulo no membranal que no está presente en las células vegetales y que es definido como el
centro organizador de los Microtúbulos de la célula. Normalmente se localiza a un lado del núcleo, junto
a su membrana, y tiene la particularidad de que cuando la célula va a entrar en mitosis genera una copia
del centrosoma.
COMPOSICIÓN
Centriolos: Dos estructuras de forma cilíndrica dispuestas
en ángulo recto y que reciben el nombre de Citocentro o
Diplosoma. Cada centriolo está formado por 9 tripletes
interconectados de microtúbulos, compuestos de Tubulina.
Aster: Conjunto de microtúbulos que salen desde el centro
del centrosoma y se alargan hacia fuera.
Matriz del centrosoma: Región del citoplasma que rodea los
centriolos y que está compuesta por una red de pequeñas
fibras, de composición exacta desconocida (sólo se sabe que
hay tubulina).
FUNCIÓN
Se divide en la división celular formando dos centriolos
iguales y perpendiculares a los anteriores, emigrando a los
polos celulares cada pareja de centriolos, formando así el
huso acromático por condensación de las proteínas
globulares que se encuentran en el citoplasma.
Los centriolos, por su parte, son estructuras que también
participan en la formación de Cilios y Flagelos, ya que en el
origen de éstos se encuentra una estructura denominada
Corpúsculo Basal que es interconvertible con el centriolo.
CILIOS Y FLAGELOS
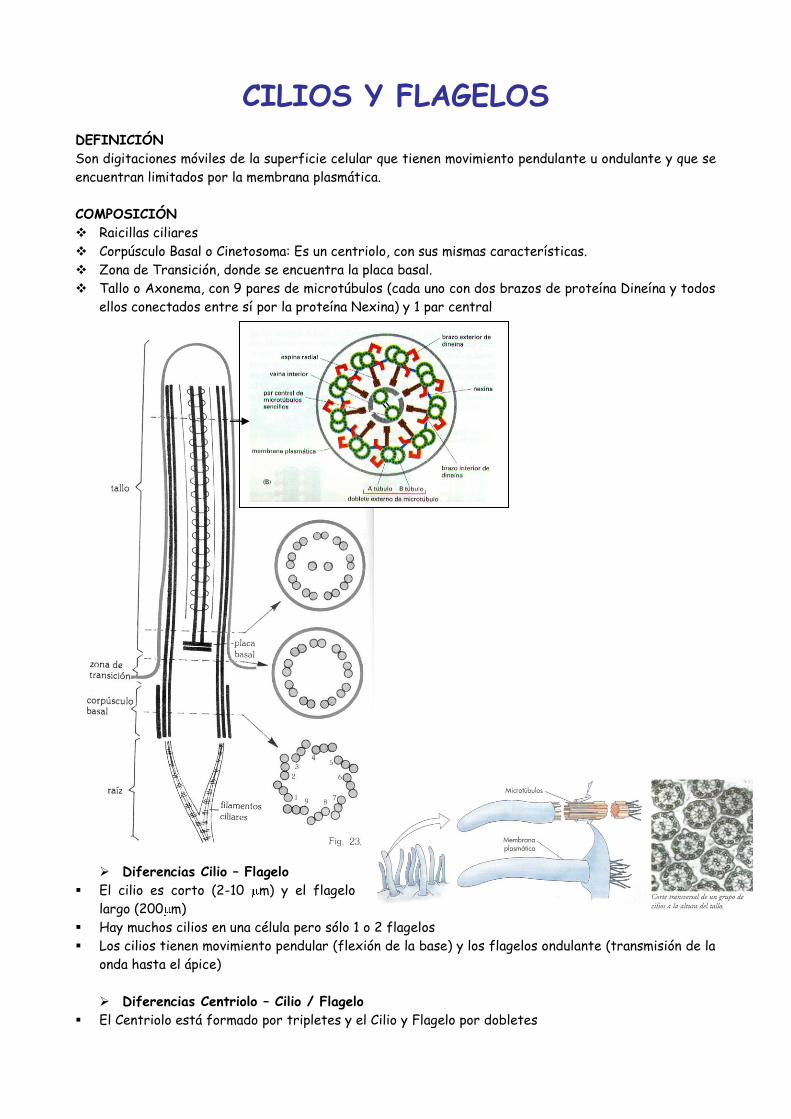
DEFINICIÓN
Son digitaciones móviles de la superficie celular que tienen movimiento pendulante u ondulante y que se
encuentran limitados por la membrana plasmática.
COMPOSICIÓN
Raicillas ciliares
Corpúsculo Basal o Cinetosoma: Es un centriolo, con sus mismas características.
Zona de Transición, donde se encuentra la placa basal.
Tallo o Axonema, con 9 pares de microtúbulos (cada uno con dos brazos de proteína Dineína y todos
ellos conectados entre sí por la proteína Nexina) y 1 par central
Diferencias Cilio – Flagelo
El cilio es corto (2-10 m) y el flagelo
largo (200 m)
Hay muchos cilios en una célula pero sólo 1 o 2 flagelos
Los cilios tienen movimiento pendular (flexión de la base) y los flagelos ondulante (transmisión de la
onda hasta el ápice)
Diferencias Centriolo – Cilio / Flagelo
El Centriolo está formado por tripletes y el Cilio y Flagelo por dobletes

VACUOLAS
INCLUSIONES CITOPLASMÁTICAS
Son vacuolas que presentan en su interior sustancias no acuosas
ORGÁNULOS CELULARES

Se encargan de dirigir el proceso de traducción o biosíntesis de
proteínas. Para ello, la subunidad menor se une al ARNm, que será
leído y cuya información será traducida de un lenguaje escrito bajo
la forma de ribonucleótidos a un lenguaje en forma de aminoácidos,
para incorporarse, algo más tarde, la subunidad mayor.
Es muy típico que una misma molécula de ARNm sea leída de forma
secuencial por varios ribosomas (entre 5 y 40) de forma que, debido
a la longitud progresivamente menor de las cadenas polipeptídicas en
síntesis, el conjunto, conocido como polirribosoma o polisoma, adopta
un aspecto característico en forma de "árbol de navidad".
RIBOSOMAS
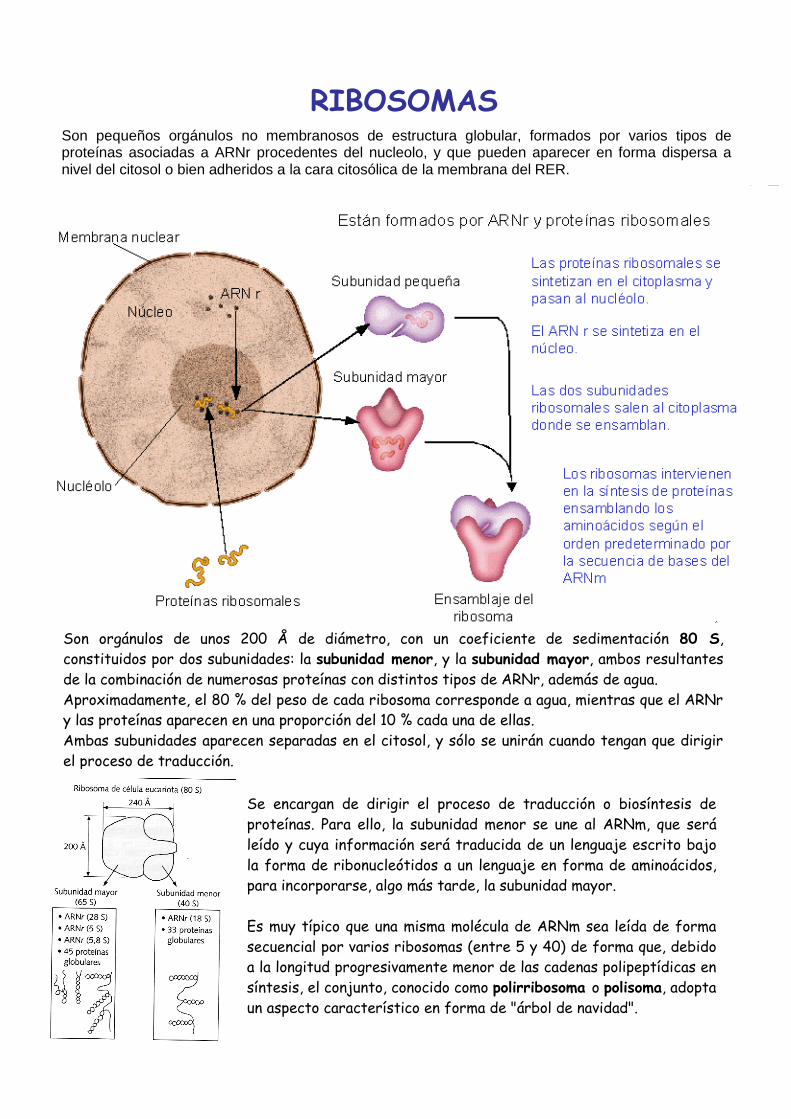
Son pequeños orgánulos no membranosos de estructura globular, formados por varios tipos de proteínas asociadas a ARNr procedentes del nucleolo, y que pueden aparecer en forma dispersa a nivel del citosol o bien adheridos a la cara citosólica de la membrana del RER.
Son orgánulos de unos 200 Å de diámetro, con un coeficiente de sedimentación 80 S,
constituidos por dos subunidades: la subunidad menor, y la subunidad mayor, ambos resultantes
de la combinación de numerosas proteínas con distintos tipos de ARNr, además de agua.
Aproximadamente, el 80 % del peso de cada ribosoma corresponde a agua, mientras que el ARNr
y las proteínas aparecen en una proporción del 10 % cada una de ellas.
Ambas subunidades aparecen separadas en el citosol, y sólo se unirán cuando tengan que dirigir
el proceso de traducción.


















