
1. INTRODUCCIÓN -...
84
1. INTRODUCCIÓN La producción de frutales tipo nuez en Chile está atravesando por una fase nueva de desarrollo. Ha dejado de ser una alternativa agronómica de bajo interés económico, para ubicarse competitivamente en zonas de privilegio en el área de plantaciones frutícolas de exportación del país. Introducción de variedades y especies, en conjunto con oportunidades comerciales en el plano internacional, son las razones del nuevo interés que se vive en el país. EL nogal se presenta actualmente como la principal especie de frutos de nuez, alcanzando una superficie de 7.700 hectáreas según el último censo de 1997, de las cuales 5.673 se encuentran en producción y 1.805 están en formación, concentradas principalmente entre la V y VI Región representando el 91% de la superficie nacional (ODEPA, 1998). Más de la cuarta parte de la superficie actualmente plantada corresponde a huertos injertados, destacando al cultivar Serr como el mayoritario. El nogal común (persa, inglés) corresponde a la especie Juglans regia, perteneciente al género Juglans, familia de las Juglandáceas, del orden Juglandales (LUNA, 1990). En la actualidad, se estima que los nocedales chilenos proceden todavía, en una alta proporción, de plantas de semilla. Aún cuando es difícil proporcionar cifras exactas de la composición varietal, ésta ha cambiado rápidamente en los últimos años, llegando según estimaciones no oficiales, al 50% del total de la superficie plantada con árboles injertados. Tanto así, que nuevas empresas con superficies de más de 100 hectáreas comienzan a desarrollarse desde fines de la década de los ‘90, dado que la mecanización de las labores en la actividad va a la cabeza de las innovaciones de esta especie (LEMUS, VALENZUELA y LOBATO, 2001).
Transcript of 1. INTRODUCCIÓN -...

1. INTRODUCCIÓN
La producción de frutales tipo nuez en Chile está atravesando por una fase nueva de
desarrollo. Ha dejado de ser una alternativa agronómica de bajo interés económico,
para ubicarse competitivamente en zonas de privilegio en el área de plantaciones
frutícolas de exportación del país. Introducción de variedades y especies, en conjunto
con oportunidades comerciales en el plano internacional, son las razones del nuevo
interés que se vive en el país.
EL nogal se presenta actualmente como la principal especie de frutos de nuez,
alcanzando una superficie de 7.700 hectáreas según el último censo de 1997, de las
cuales 5.673 se encuentran en producción y 1.805 están en formación, concentradas
principalmente entre la V y VI Región representando el 91% de la superficie nacional
(ODEPA, 1998). Más de la cuarta parte de la superficie actualmente plantada
corresponde a huertos injertados, destacando al cultivar Serr como el mayoritario.
El nogal común (persa, inglés) corresponde a la especie Juglans regia, perteneciente
al género Juglans, familia de las Juglandáceas, del orden Juglandales (LUNA, 1990).
En la actualidad, se estima que los nocedales chilenos proceden todavía, en una alta
proporción, de plantas de semilla. Aún cuando es difícil proporcionar cifras exactas
de la composición varietal, ésta ha cambiado rápidamente en los últimos años,
llegando según estimaciones no oficiales, al 50% del total de la superficie plantada
con árboles injertados. Tanto así, que nuevas empresas con superficies de más de 100
hectáreas comienzan a desarrollarse desde fines de la década de los ‘90, dado que la
mecanización de las labores en la actividad va a la cabeza de las innovaciones de esta
especie (LEMUS, VALENZUELA y LOBATO, 2001).

Serr ha sido una de las variedades más plantadas en California, sin embargo, en la
década de los 80´ se comenzó a detectar ciertos desórdenes productivos que
provocaba un descenso brusco del rendimiento. En California, donde se produce el
99% de la producción de los EEUU, la abscisión de flores pistiladas (AFP) ha
emergido como un serio problema en algunos cultivares de nogal (CATLIN et. al,
1987). La variedad Serr, la que tiene una pérdida de flores a causa de la AFP mayor a
90% aparece como el cultivar más susceptible.
Hasta ahora los investigadores de la Universidad de California, Estados Unidos, han
dirigido su atención a que la posibilidad del aborto de flores pistiladas pueda ser
causado por un exceso de polen en estas flores, descartando posibles causas como
bajos niveles de nitrógeno, competencia por los carbohidratos, excesivo
sombreamiento; por otro lado, no se ha podido establecer una relación de la AFP con
factores ambientales o culturales como edad de la planta, competencia entre órganos
de ésta, sistema de poda, alternancia de carga, plagas y enfermedades (RYUGO et al.,
1985; CATLIN et al., 1987).
Los primeros resultados de los experimentos con flores que recibieron altas dosis de
polen fueron promisorios; el aborto de flores pistiladas fue directamente proporcional
a la carga de polen, los cultivares de nogal tuvieron variadas respuestas a las altas
dosis de polen, y el aborto de flores pistiladas se presenta sin importar la fuente de
polen (POLITO et al., 1998).
Considerando que la superficie de nogales del cultivar Serr en Chile ha aumentado en
la última década (VALENZUELA, LOBATO y LEMUS, 2001), resulta importante
estudiar el comportamiento de este cultivar frente al aborto de flores pistiladas, bajo
las condiciones locales y aún más, el comportamiento del mismo huerto en un
segundo año de medición.

De acuerdo a lo anterior, se plantea que si el cultivar Serr manifiesta aborto de flores
pistiladas debido a un exceso de polen, produciendo una baja en la productividad, el
manejo de la polinización mediante la remoción de amentos (flores masculinas),
logrará un aumento significativo en los rendimientos.
El objetivo global de este estudio es dimensionar la pérdida de productividad en el
aborto de flores pistiladas, y proponer una solución.
Los objetivos específicos de este trabajo son:
- Determinar la fenología floral del cultivar Serr en la localidad de Hijuelas en la V
región.
- Determinar la efectividad del tratamiento de agitación sobre la caída de amentos.
- Cuantificar el aborto de flores pistiladas en el cultivar Serr.

2. REVISIÓN BIBLIOGRÁFICA
2.1. Antecedentes económicos del nogal :
La producción nacional de nueces es de 11.340 ton, en una superficie de 7.769 ha
(ODEPA, 2001).
La cifra de producción obtenida ha ido aumentando en nuestro país, producto de una
mayor planificación y mejor manejo de los huertos, debido a que se plantan a una
mayor densidad y se utilizan plantas injertadas (CAMPOS y VALDERRAMA,
2001).
De acuerdo a CAMPOS y VALDERRAMA (2001), la producción de nueces se
destina principalmente al mercado de exportación. Las exportaciones de la fruta es
con o sin cáscara, pero en el último período ha aumentado la exportación de la fruta
descascarada debido al cambio de los hábitos de consumo.
Nuestro país exporta nueces a unos 20 países, sin embargo, existe una alta
concentración de la exportación en dos países, Argentina y Brasil, que representan
más del 70% de los envíos de nueces que efectúa el país.
2.2. Antecedentes de la especie :
Juglans regia es originaria de países asiáticos, desde los Cárpatos hasta Afganistán,
sur de Rusia y norte de India. Procedente de la región del Himalaya, Persia, llegó
hasta Italia donde obtuvo su nombre Lovis glans (bellota de Júpiter) y del latín Dios
bálanos (Castaña de Zeus) (PASTORE, 2002).

2.2.1. Características morfológicas
Árbol muy vigoroso que puede llegar a alcanzar 27 m de altura. Su copa es ramosa,
extendida, de forma esférica comprimida. El tronco es derecho, cubierto con una
corteza cenicienta y gruesa, que en las ramas jóvenes es lisa y de color rojo oscuro,
mientras que en las ramas viejas es agrietada y parda (PASTORE, 2003).
El sistema radicular es bastante desarrollado, formado por una raíz principal pivotante
y un sistema secundario de raíces someras y robustas. Las raíces se ubican
notablemente extendidas, tanto en sentido horizontal como vertical. CATLIN (1998)
señala que en suelos de textura franca, bien drenados, lo normal es que la exploración
radical no sobrepase los 2 metros y un alto porcentaje de ellas se encuentre en el
primer perfil del suelo.
Según MUNCHARAZ (2001) las yemas presentes en el árbol pueden ser de
vegetativas o yemas fructíferas y a su vez de distinta naturaleza, encontrando las
siguientes:
- Yemas de madera sencilla. Sólo hay una yema principal claramente diferenciada,
a veces acompañada por una inflorescencia estaminada.
- Yemas de madera doble. Están compuestas por una yema principal más
desarrollada y otra secundaria situada por debajo de la principal. Estas yemas
pueden ser folíferas, floríferas o mixtas.
- Yemas floríferas masculinas. Aparecen sustituyendo a una o a las dos yemas
dobles vistas anteriormente, de forma que de ellas pueden formarse dos amentos,
un brote y un amento o dos brotes.
- Yema florífera femenina. Generalmente se sitúan en posición terminal, sobre
brotes formados en el mismo periodo vegetativo.

2.2.2. Polinización y fecundación
El nogal es una especie monoica, presenta las flores masculinas y femeninas
separadas, pero situadas sobre el mismo pie (ALETA y NINOT, 1993). Son
genéticamente auto e intercompatibles, pero dentro de cultivares individuales el
periodo de oferta del polen de las flores masculinas no traslapa completamente el
periodo de receptividad de las flores femeninas, fenómeno conocido como dicogamia
(POLITO, 1998).
El término dicogamia es usado para referirse a los eventos que ocurren dentro de las
flores donde los órganos masculino y femenino maduran en momentos distintos,
dentro de una planta individual, donde la floración estaminada y pistilada están
separadas temporalmente (POLITO Y PINNEY, 1997).
POLITO (1998) señala que el nogal presenta dicogamia y existen dos formas de este
comportamiento entre cultivares. La mayoría de aquellos plantados hoy en día
presentan dicogamia protoandria, es decir, las flores masculinas entregan el polen
antes que las flores pistiladas sean receptivas. Unos pocos cultivares (Chico, Amigo),
son protogíneos; en este caso, la receptividad de las flores femeninas ocurre antes que
la entrega de polen.
La dicogamia protoandria es favorecida por el clima cálido y la dicogamia protoginea
por el clima frío, por lo que en esta especie se presenta una heterodicogamia.
Hereditariamente es dominante el carácter protoándrico (BINGGELI, 2003)
El nogal es una especie cuya polinización es efectuada por el viento (anemófila),
para lo cual el polen está particularmente adaptado, por ejemplo, se ha constatado la
presencia de 5 granos/cm2/12 horas a una distancia de 160 m del emisor (GIL, 2000).

Las flores también están adaptadas a este tipo de polinización. MUNCHARAZ
(2001) afirma que las flores femeninas son apétalas y con una gran superficie
estigmática, preparada para recibir los ligeros granos de polen que llegan al estigma.
Las flores pistiladas solamente son receptivas por un tiempo limitado; si no son
polinizadas durante ese período no producirán fruta. La receptividad se puede juzgar
por apariencia visual directa de los estigmas. Se inicia cuando los dos lóbulos del
estigma comienzan a separarse uno del otro y a crecer . Cuando los lóbulos de los
estigmas se han desarrollado y forman un ángulo de 45º entre ellos, termina su
periodo de receptividad hasta finalizar cuando los lóbulos se hunden y adquieren
manchas pardas (POLITO, 1998). El período receptivo de una flor femenina es de
entre tres y seis días, dependiendo de si el clima es cálido y seco o fresco y húmedo.
La floración femenina puede durar hasta 15 días y la masculina hasta 10 días (GIL,
2000).
El polen es de corta vida después que es liberado desde el amento, siendo viable entre
4-24 horas (POLITO, 1985). Una vez depositado en el estigma receptivo pasa una
semana hasta que ocurre la fecundación y siete a diez días más tarde el cigoto
unicelular inicia su división; en este caso, los estigmas se tornan pardos y secos
mientras que en las flores no fertilizadas se mantiene el aspecto original (GIL, 2000).
Las flores que no han sido polinizadas continuarán su crecimiento al menos durante
tres semanas desde la finalización del periodo de receptividad. Dicho crecimiento es
similar al presentado por las flores polinizadas. Después de este periodo las flores no
polinizadas caerán (POLITO, 1998).
La polinización es necesaria para la fecundación y para el desarrollo del embrión
comestible (FORDE y GRIGGS, 1972). Solamente en muy pocas variedades como
Geisenheim 139 de Alemania y Esterhazy II de Hungría, existe apomixis en algunas
ocasiones (GERMAIN, JALINAT y MARCHOU, 1975)

2.2.3. Tipos de fructificación
Según MUNCHARAZ (2001), los tipos de fructificación en nogal están ligados con
la entrada en producción y la productividad. La entrada en producción está
directamente ligada a la aparición de las flores masculinas, que en la mayoría de las
variedades aparece uno o dos años más tarde que las femeninas. La capacidad
productiva va íntimamente ligada a una fructificación lateral, definida por la
capacidad de producción de flores femeninas en el extremo de brotes nacidos de
yemas laterales de crecimientos de un año (CHARLOT y GERMAIN, 1988).
De acuerdo a lo anterior, las variedades se pueden clasificar de acuerdo a los
siguientes tipos de fructificación:
• Fructificación apical y ramificación débil: Tiene gran vigor y una entrada en
fructificación tardía, ya que sólo fructifican en el extremo de la rama. Su
productividad es débil.
• Fructificación apical y ramificación fuerte: Tiene un comportamiento intermedio.
• Fructificación sobre brindillas laterales: Produce más y entran antes en
producción; consecuentemente su vigor es menor, debido a la competencia precoz
que ofrecen los numerosos frutos.
2.2.4. Morfología de las flores y frutos
Flores masculinas o estaminadas: son pequeñas, individuales y relativamente
insconspicuas. Se encuentran agrupadas en racimos cilíndricos colgantes
denominados amentos, los cuales alcanzan una longitud de 8 a 10 cm. Las flores se
elongan tardíamente en la temporada previa al crecimiento. Los amentos se
encuentran lateralmente en las ramillas de un año de edad y se forman tres a seis

semanas después de la brotación (POLITO, 1998). De acuerdo a MUNCHARAZ
(2001), las flores individuales carecen de pétalos y consisten en sépalos verdes.
Cada flor contiene unos 40 estambres, los que pueden contener cientos de granos
microscópicos de polen. Cuando las condiciones climáticas son adecuadas, las anteras
se abren y liberan el polen que transportado por el viento, alcanza el estigma, que es
la parte receptiva de la flor femenina. En el estigma se producen condiciones
adecuadas de humedad (su falta podría secar el grano y de esta forma anular su poder
germinativo), para que el polen germine. La germinación produce la formación de un
tubo polínico que emerge a través de los poros del polen. En el extremo del tubo van
las células germinales
masculinas, el esperma, que posteriormente se unirá con la célula germinal femenina
(óvulo) (MUNCHARAZ, 2001).
Aunque muchos granos de polen pueden llegar al estigma y se pueden formar muchos
tubos polínicos, solamente uno (o ninguno), alcanzará el óvulo. Para llegar al óvulo,
el tubo polínico tiene que atravesar todo el pistilo.
El tiempo transcurrido entre la llegada del polen al estigma y la fecundación puede
ser de una semana (POLITO, 1998).
Experimentos conducidos por McGRANAHAN et al. (1994) indican que el déficit de
agua en los pistilos puede jugar un rol importante creado por un mal balance entre la
transpiración y el estatus hídrico de la raíces, lo que traerá como consecuencia una
deshidratación de los pistilos y una posterior caída de éstos.
Después de numerosas investigaciones hechas por diversas personas, se ha llegado a
la conclusión de que al tener una alta presencia de granos de polen sobre el estigma,
se produce un aborto de frutos recién cuajados. Así, KRUEGER (2000) concluye que
el conteo de granos de polen sobre las flores pistiladas muestra un alto porcentaje de

aborto de flores siempre asociado con un alto número de granos de polen. Las curvas
de respuesta a la dosis indicaron que 70 a 100 granos de polen por flor pistilada
inducirían un aborto del orden del 50%. Esta relación fue consistente para huertos de
diferentes partes de California.
Análisis hechos por McGRANAHAM et al. (1994) demostraron diferencias
significativas entre pistilos con mayor o menor presencia de polen. De esta manera,
pistilos que tenían entre 130 y 180 granos de polen no presentaban problemas, ahora
bien, si esta cifra se elevara entre 230 y 320 existirían problemas de aborto floral.
CATLIN et al. (1987) proponen que alguna influencia dentro del pistilo causó un
daño celular que detuvo el crecimiento e inició el proceso que lleva a la abscisión 10
a 14 días más tarde. Los eventos que llevan al daño parecen ocurrir en un período
cercano a la detención del crecimiento, es decir, uno o dos días antes. Por lo tanto, el
posible aborto de flores no parece ser una expresión de defectos inherentes del tejido
del ovario. Luego de hacer un examen histológico se evidencia que las células de la
pared interior del ovario (tegumentos o integumentos), y las evaginaciones que
rodean los óvulos se desintegran, presentándose éstas y los óvulos de un color negro
y necróticos; lo cual sugiere que el daño es dentro del pistilo y no por causas externas.
Flores femeninas o pistiladas: La iniciación de las flores ocurre durante la
temporada de crecimiento previa a la floración y las etapas finales del desarrollo de la
flor ocurren en la primavera, durante las semanas anteriores a la floración (LIN,
SHABNAY y RAMOS 1977; POLITO, 1988; POLITO y LI 1985).
Las flores pistiladas se forman frecuentemente en pares y se desarrollan en una yema
terminal en todos los cultivares (fructificación terminal), y en yemas laterales en
ciertos cultivares (fructificación lateral) (KRUEGER, 2000).

De acuerdo a POLITO (1998), la parte exterior de la flor está compuesta por el
involucro, las brácteas y cuatro sépalos. Todos ellos formarán, en la madurez, el
pericarpio de la nuez. La parte externa del involucro está formada por una sustancia
pegajosa de naturaleza cerosa, segregada por glándulas pilosas superficiales.
Rol importante juega el estigma, ya que si bien está adaptado para capturar el polen
llevado por el viento, también lo debe reconocer y rechazar si es que fuera de otra
especie. El estigma tiene dos brazos plumosos. Las células del estigma segregan un
líquido que forma una delgada cubierta superficial. Esta exudación facilita la
hidratación de los granos de polen que se pudieran depositar y por lo tanto, hacen
posible la germinación del polen y el crecimiento inicial del fruto (POLITO, 1998).
Con respecto al estilo, éste es muy corto y une el estigma con el ovario. En el ovario
hay
una cavidad central (lóculo), que se divide en cuatro partes, por dos paredes
denominadas tabiques mayor y menor. Debajo del lóculo hay un ovario simple que
dará lugar al grano o semilla después de realizada la fecundación y el cuajado, y
donde el tegumento será la película que lo cubrirá (piel del grano). El saco
embrionario contenido en la nucela es una estructura altamente organizada,
compuesta por siete células, una de las cuales es la célula huevo, que formará el
cigoto al unirse con la célula sexual masculina.
Fruto: El desarrollo del fruto comienza al momento de la polinización y la
fertilización de la flor pistilada. El fruto comienza a crecer cuando los tubos polínicos
están creciendo en el estigma. La unión de las células sexuales masculinas y
femeninas origina el cigoto (POLITO, 1998).
La célula sexual femenina se encuentra dentro del óvulo (futuro embrión del fruto),
que está formado por distintas partes:
- El tegumento, que es una pequeña envoltura del óvulo.

- La nucela es la parte del óvulo que da volumen a éste durante la fecundación.
Desaparece en la madurez.
- El saco embrionario está formado por siete células, una de las cuales es la célula
huevo o célula sexual femenina, que se unirá con una de las dos células
espermáticas masculinas para formar el cigoto.
Luego de la fecundación del óvulo, existen una serie de fases o períodos en donde el
fruto comienza su crecimiento, destacando fundamentalmente: multiplicación celular,
engrosamiento celular y maduración (GIL-ALBERT, 1996).
Según GIL (2000), la primera etapa de crecimiento dura 7-8 semanas y pocas
semanas
después el involucro y el endocarpio alcanzan su tamaño máximo por aumento del
número y tamaño de células. La etapa final de crecimiento del fruto y del embrión se
debe a cambios químicos y aumento de materia seca.
El tiempo de los estados más importantes en el desarrollo del fruto del nogal y la
curva de crecimiento se muestran en el Anexo 1.
La carne y el fruto entero experimentan un breve de crecimiento relativamente lento
durante la mitad de la temporada. En el fruto la disminución del crecimiento
comienza cerca de las ocho semanas después de floración y continúa por tres a cuatro
semanas; corresponde al período de rápida expansión de los cotiledones y
endurecimiento de la cáscara. Subsecuentemente, el crecimiento de la carne
disminuye por un tiempo, pero la rápida proporción de aumento de peso, se reanuda
mientras el embrión acumula materia seca. El tamaño final de la fruta es alcanzado
relativamente rápido. La disminución de peso fresco hacia la cosecha es atribuible a
la pérdida de agua, particularmente en el pelón (POLITO, 1998).

El fruto es una drupa globosa formada por dos valvas indehiscentes, de 4-5 cm de
largo. La nuez consta de dos gruesos tabiques y de cuatro celdas incompletas
indehiscentes o finalmente separadas en dos valvas (WESTWOOD, 1993). El
mesocarpo, que es verde y ennegrece rápidamente, encierra un hueso (la ‘nuez’),
formado por el endocarpo leñoso, debido a su contenido de lignina; y la semilla, con
sus característicos cotiledones muy lobulados, casi cerebriformes. Esta última es la
parte comestible (RENOBALES, 2001).
2.2.4.1. Estados fenológicos de las flores del nogal
EL conocimiento de los estados de desarrollo de las plantas y la existencia de una
nomenclatura para su identificación, son herramientas necesarias para su
combinación con algunas técnicas de cultivo, tales como programación de la
polinización, si existe o no concordancia entre cultivares, aplicaciones de pesticidas
y/o fungicidas, entre otros. De esta manera, se entiende que la fenología anatómica y
morfológica de la diferenciación floral juega un papel importante al proporcionar
datos que ayudan a programar prácticas culturales que puedan influir en la época e
intensidad de la floración.
POLITO (1998) destaca la importancia de predecir la extensión de traslape de la
floración masculina y femenina para determinar la factibilidad de remover amentos.
Hasta el momento, esto se puede hacer solamente por medio de una observación
cuidadosa del progreso del ciclo de floración.
MUNCHARAZ (2001) agrega que el valor de utilizar un lenguaje internacional,
obliga a aceptar los estados fenológicos florales tipo, ya determinados y más
corrientemente aceptados. De acuerdo a lo anterior, el autor establece:
Flor femenina y hojas
Af: Yema en reposo. De esta forma pasa el invierno, recubierta por escamas.

Af2: Caída de escamas duras. La yema queda protegida por otras escamas poco
diferenciadas y semi-membranosas.
Bf: Botón blanco o lanoso. La yema se hincha y aparecen las brácteas subyacentes
recubiertas de una pelusa blanquecina.
Cf: Desborre. Aparecen nuevas brácteas y los foliolos terminales de las hojas
exteriores.
Cf2: Primeras hojas. La yema se abre y se separan las escamas, brácteas y foliolos.
Df: Separación de las hojas. La yema está totalmente abierta y las primeras hojas se
despliegan y separan.
Df2: Principio de foliación. Las primeras hojas están completamente abiertas y
erguidas, toman una posición oblicua y dejan aparecer las flores femeninas.
Ef: Aparición de las flores femeninas.
Ff1: Aparecen los estigmas, que empiezan a separarse y pasan de rojo a verde
amarillento. En este estado comienza su receptividad, y llega a su punto máximo,
mostrando los estigmas bien expandidos y sus dos lóbulos están visiblemente
separados.
Ff2: Plena floración femenina. Los estigmas se abren y finalizan su receptividad. Su
color es verde pálido.
Gf: Marchitamiento de los estigmas. Se desecan, pasando del marrón al negro, ya no
son receptivos.
I: Cuajado
J: Engrosamiento del fruto.
K: Lignificación de la cáscara. La cáscara está completamente formada. 7
Di: Agostamiento de la madera. Los limbos de las hojas han acabado su crecimiento
L: La corteza del fruto se agrieta
Lo: Se puede ver la nuez en el interior
Dj: Amarillamiento de las hojas
Mo: La corteza se encuentra completamente dehiscente
O: Caída de la nuez

Dz: Caída de las hojas
Inflorescencia masculina
Amr: El amento se diferencia en verano, adquiriendo un tinte rosado.
Amv: El amento crece ligeramente, alcanzando 0,5 cm. Toma color verde
Amg: El amento deja de crecer para invernar. Toma color grisáceo
Bm: Se reanuda el crecimiento del amento después de pasado el invierno. Comienzan
a formarse los granos de polen. La longitud del amento puede llegar a 2 cm
Cm: El amento alcanza el grosor de un lápiz. Color verde claro. Los glomérulos de
las flores se distinguen bien. Un glomérulo es la estructura donde se encuentra cada
flor de la inflorescencia.
Dm: El amento pierde rigidez y se separan los glomérulos
Dm2: El amento cuelga, consecuencia de la separación de los glomérulos.
Em: Los glomérulos están completamente abiertos. Las anteras comienzan a
separase.
Em2: Las anteras finalizan su separación y comienzan a amarillear.
Fm: Total amarillamiento y comienzo de la dehiscencia de las anteras desde la base
del amento.
Fm2: Dehiscencia total de las anteras y plena emisión de polen.
Gm: Las anteras, ya sin polen, toman un color pardo
Gm2: Las anteras están completamente desecadas.
Hm: El amento completamente seco cae.
Algunos estados fenológicos florales masculino y femenino se presentan en las
figuras 1 y 2 respectivamente.

2.2.5. Requerimientos climáticos
Para el caso de las variedades californianas como Serr, las nueces de mejor calidad se
cosechan en lugares donde se presentan veranos prolongados y no se registran
temperaturas muy elevadas, de manera que no excedan los 28°C, teniendo siempre
buena humedad en el suelo, alcanzando la nuez su total desarrollo, sabor y cualidades
gustativas. La temperatura óptima para una adecuada actividad del grano de polen es
alrededor del los 25°C. Las nueces de algunos cultivares son particularmente
sensibles al calor. A mediados de verano, durante el desarrollo de la carne, las
temperaturas mayores a 30ºC pueden provocar quemaduras por sol, por lo tanto las
nueces resultarán vacías, además la semilla se puede oscurecer, deshidratarse o
incluso adherirse al interior de la cáscara, lo que reduce su valor (SIBBETT,
COATES y EDSTROM, 1998).
La gran limitante en cuanto a condiciones climáticas es la presencia de heladas
primaverales, que corresponde a la época de floración (HUDSON, WILLIAM y
ANTOM, 1981); y uno de los mayores inconvenientes que presenta el cultivo del
nogal, es su tendencia a iniciar el transporte de savia ante el primer aumento de
temperatura. Por lo tanto heladas fuera de estación serán de mayor impacto y sobre
todo aquellas que se producen al comienzo de la primavera. Las heladas tempranas
del otoño también son peligrosas, ya que desecan el extremo de las ramas poco
lignificadas, disminuyendo así la producción (CECICH, 2003).

FIGURA 1. Distintos estados florales masculinos en nogal cv. Serr. A. Amentos en estado de reposo durante julio 2002. B. Amentos empezando a elongar a comienzos de brotación, finales de agosto 2002. C. Amentos completamente elongados, finales de septiembre 2002.
A BC

FIGURA 2. Secuencia de flor pistilada durante la temporada 2002. A. La ramilla se ha expandido y aparecen las primeras flores femeninas. B y C. Los estigmas se muestran el color rojizo – amarillento, típico de un estado de receptividad. D. Los estigmas se muestran totalmente expandidos, ya no son receptivos.
A
B
C D

El resultado de estas heladas tempranas o tardías, son muy perjudiciales para la
fructificación, ya que las nueces se originan generalmente en brotes de yemas
terminales, no alterando todo el vigor del nogal, a no ser que sean muy intensas y
frecuentes.
En invierno pueden producirse algunos daños en las ramas y yemas del árbol, si la
temperatura cae por debajo de –7ºC. Si en la zona las temperaturas de invierno llegan
con normalidad a –9ºC, no es recomendable la plantación de nogales (SIBBETT,
COATES y EDSTROM, 1998).
La mayoría de los cultivares de nogal requieren aproximadamente 800 horas con
temperaturas bajo los 7ºC durante el invierno para que los árboles broten y produzcan
normalmente (SIBBETT, COATES y EDSTROM, 1998). Las variedades francesas
pueden llegar a necesitar 1500 horas frío, frente a las 300 de algunas variedades de
California (MUNCHARAZ, 2001).
Se ha visto que en lugares donde la oferta hídrica no sobrepasa los 500 mm anuales,
el nogal se ha desarrollado normalmente. No sólo es importante la cantidad total de
precipitaciones, sino también su distribución anual; si las lluvias son deficientes y
mal distribuídas se deberá recurrir al riego como necesidad para obtener buenas
producciones (CECICH, 2003).
2.2.6. Actuales recomendaciones sobre el uso de polinizadores
Entre los asuntos más apremiantes de las actuales investigaciones acerca del papel
que juega el polen en la AFP se encuentra en la necesidad de reevaluar el rol del
polinizador en los huertos de nogal (POLITO, 1998).

El polen es requerido para la cuaja del cultivo, pero no está claro la necesidad y
densidad de los polinizantes (KRUEGUER, 2000).
Según POLITO (1998), hay evidencia que sugiere que las plantaciones sin
polinizante están sujetas a una caída más tardía de las flores, probablemente debido a
una inadecuada polinización.
El número de árboles de la variedad polinizadora a intercalar en la plantación,
independiente de la variedad comercial, debido a que el polen del nogal es
intercompatible, es un tema aún no resuelto (MUNCHARAZ, 2001). La proporción
de árboles del polinizador puede variar entre un 2.5 y 10% (SIBBETT, COATES y
EDSTROM, 1998).
FORDE y GRIGGS (1975) sugieren que cerca de un 10% de los árboles en una
plantación sea de un cultivar con un buen traslape de la floración estaminada con la
floración pistilada del cultivar principal.
Los siguientes aspectos deben considerarse al momento de planificar la plantación
(MUNCHARAZ, 2001):
- La ubicación de los polinizadores debe considerar la recolección, teniendo en
cuenta que la madurez de la variedad base y la polinizante probablemente no
coincidan.
- Los árboles polinizadores se deben ubicar perpendicularmente a la dirección
dominante de los vientos, debido a su polinización anemófila, a intervalos
regulares, y espaciados cada 100 – 200 metros como máximo.
- Los árboles polinizadores se deben colocar en posiciones permanentes de manera
de no eliminarlos inadvertidamente cuando sea el momento de eliminar los
árboles intercalados.

El problema que ha surgido con la variedad Serr, y del que también participan
cultivares tales como Vina, Chico y Chandler, es el denominado abscisión de flores
pistiladas (AFP), producido por un exceso de polen en la plantación llegando a
disminuir las cosechas hasta en un 90% (CATLIN y OLSSON, 1990). Por estos
motivos, el cv. Serr se está plantando sin árboles polinizadores. En los huertos
antiguos se está eliminando los árboles de la variedad polinizadora; y las para las
demás variedades se recomienda plantar solamente con un 5% - 10% según decrece
su susceptibilidad al aborto (GIL, 2000).
Ciertamente, la recomendación según POLITO (1998) para los árboles polinizadores
ha disminuido en años recientes y actualmente es de 2 a 3% dependiendo de los
factores que se mencionan a continuación:
- Traslape entre la floración pistilada y estaminada en el cultivar principal.
- Susceptibilidad del cultivar principal al AFP.
- Densidad de nogales en el área, debido a su polinización por el viento.
- Experiencia local con la polinización del nogal
2.2.7. Manejo de la carga de polen en huertos de Serr
El polen en un huerto Serr tiene dos fuentes primarias: el cultivar polinizador y los
árboles Serr. La significancia del propio polen Serr varía de año en año, dependiendo
de la extensión del traslape entre los períodos de floración femenina y masculina.
Cuando el traslape es extenso, el polen de Serr puede ser un factor importante en el
manejo de la carga total de polen sobre el umbral de AFP y aumento de AFP
(POLITO, 1998).
Las variedades de abundante fructificación, como Serr, deben establecerse en huertos
donde la carga de polen sea muy reducida (RYUGO et al, 1985).

POLITO (1998) plantea dos formas de reducir el polen en un huerto Serr:
- Remoción de árboles polinizadores : En experimentos donde se removió Tehama
de porciones de un huerto Serr, se aumentó la producción desde 20% a 80% por
sobre las partes del huerto donde no se eliminaron los polinizantes. Sin embargo,
la remoción de polinizantes puede ser aconsejable en las siguientes situaciones: en
huertos severamente afectados por pérdidas debidas a la AFP, en huertos jóvenes
donde los polinizantes pueden ser injertados con la variedad principal, y en
huertos antiguos donde la producción del polinizante es baja.
- Remoción de los amentos del árbol polinizador y/o de la variedad comercial : Al
agitar los árboles polinizantes Tehama la AFP disminuye y los rendimientos
aumentan de entre un 20% y un 80% por sobre los controles.
La remoción de amentos puede ser manualmente (RYUGO, 1986) o por causas
naturales como daño por frío (POR y POR, 1990), la caída de amentos implica una
menor cantidad de polen disponible para la polinización (Mc GRANAHAN et al.,
1994).
2.3. Características de la variedad Serr :
Variedad originada en la Universidad de DavisCalifornia, EEUU, por E.F. Serr y H. I.
Forde en 1958. Corresponde a un cruzamiento de Payne por PI 159-568 y
seleccionado en 1963. Fue introducido a Chile en 1968 (VALENZUELA,
ESPINOZA y PARRA, 1992).
Las características más destacables de esta variedad son su elevada capacidad
productiva, pese a tener solo el 50% de brotes fructíferos ubicados en posición lateral,

y su gran vigor, que facilita tanto su formación como el mantenimiento del equilibrio
productivo en plantaciones adultas (ALETA y NINOT, 1993).
HENDRICKS et al., (1998) indican que la fecha de brotación de Serr es a mediados o
fin de marzo (California, Hemisferio Norte) es decir, casi la misma o un día antes que
Payne. La nuez de Serr es grande, la carne pesa alrededor de 7,8 gramos. La nuez es
ovalada y lisa con sello de cáscara regular a bueno. El color es muy bueno, el 70 a
80% de las nueces son clasificadas como color claro. El porcentaje de carne es alto,
57%. La cosecha de Serr es temprana. Esta variedad tiene muchos atributos, muy
deseables, por lo que la demanda del mercado por ella es fuerte. Serr puede ser una
sabia decisión para un productor en un área donde los problemas de peste negra
usualmente no son severos y donde se han alcanzado altos rendimientos. Los árboles
de Serr son grandes, la forma del árbol es moderadamente extendida y el vigor es
bueno a excesivo. Debido a su temprana brotación, Serr es susceptible a la polilla del
manzano y a la peste negra.
CATLIN y POLITO (1989), indican que Serr ha sido una de las variedades mas
plantadas en California y en el resto de los países donde se cultiva el nogal, sin
embargo, a partir de los años 80´ se empezó a detectar en esta variedad ciertos
desarreglos productivos que provocaban un descenso brusco de la producción. El
fenómeno se asoció a una caída prematura de flores, conocida como AFP (Aborto de
flores pistiladas) (CATLIN y POLITO, 1989), capaz de mermar las cosechas a su
quinta parte. Este fenómeno, detectado también sobre otras variedades en California,
ha encontrado en esta variedad su máxima expresión (POR y POR, 1990).

2.4. Aborto de flores pistiladas en nogal:
De acuerdo a KRUEGER (2000) el aborto de flores pistiladas se produce a principios
de temporada, cerca de dos a tres semanas después de la floración, cuando las flores
han alcanzado un tamaño de 4 a 5 milímetros.
Los pistilos parecen desarrollarse de manera normal. El alargamiento del ovario en
las flores afectadas cesa con 3 a 4 mm de diámetro. El normal curvamiento de los
estilos y estigmas cesa en una posición parcialmente completa en la etapa media y
tardía de receptividad al polen (FORDE y GRIGGS, 1975; POLITO y LI, 1985). Los
pistilos afectados permanecen unidos al brote por más de dos semanas. Cuando la
abscisión del frutito ocurre entre el pedúnculo y el eje vegetativo, todos los pistilos
(comúnmente dos) en un brote reproductivo se pierden (CATLIN y OLSSON, 1990).
Cerca de la época de abscisión, las flores del tipo AFP muestran signos de necrosis en
el ápice del estigma, en el tegumento y a través de todas las evaginaciones de la
placenta (CATLIN y POLITO, 1989).
La abscisión de flores a causa de AFP es claramente distinguible de la abscisión
causada por la falta de fertilización. Las flores no fertilizadas generalmente crecen a
más de 7 mm de diámetro y permanecen unidas por tres semanas o más (POLITO y
LI, 1985). En flores AFP, el crecimiento del pistilo cesa en un estado tardío de
receptividad del polen, indicado por la extensión del desarrollo del estigma. Los
pistilos normales, pero no polinizados, crecen mucho más antes de la abscisión que
aquellos pistilos afectados por AFP. La abscisión de los pistilos de mayor tamaño se
presenta en la zona entre el ovario y el pedúnculo, en contraste con la separación
entre el pedúnculo y el ápice vegetativo como ocurre con la AFP (HENDRICKS et
al., 1998).

2.4.1. Historia del problema de abscisión de flores pistiladas y su investigación
La pérdida de un alto número de flores pistiladas poco tiempo después de la floración
se observó primero en el cultivar Serr, luego que las primeras plantaciones
comerciales entraron en producción, a mediados de los años 70’ (POLITO, 1998).
A fines de los 80’ y principios de los 90’, los investigadores de la Universidad de
California comenzaron a investigar el rol de una carga excesiva de polen sobre la
AFP. Los primeros experimentos con flores que recibían altas dosis de polen
demostraron que la AFP estaba relacionada con la carga de polen. Los cultivares de
nogal variaban su respuesta a altas cargas de polen y la AFP no estaba relacionada
con la fuente de polen (CATLIN y OLSSON, 1990; McGRANAHAN et al. 1994).
La investigación realizada por POLITO et al. (1998) durante los 90’ en Serr demostró
que la AFP está positivamente correlacionada con la carga de polen, la distancia de la
fuente de polen, sin importar la variedad procedente y, además se ha descubierto que
reduce los rendimientos.
Se han postulado como posibles causas de la AFP bajos niveles de nitrógeno,
competencia por los carbohidratos (DENG et al., 1991), y un excesivo
sombreamiento (RYUGO et al., 1985; RYUGO, 1986).
La competencia por reservas, que comúnmente se piensa que es responsable de los
altos niveles de abscisión post-antesis de árboles frutales, no parece estar relacionada
e involucrada en la AFP (CATLIN y OLSSON, 1990). Además, CATLIN et al.
(1987) han mostrado que la AFP no está relacionada con el grado de fructificación
(índice de floración) del árbol.

Los investigadores han eliminado una larga lista de posibles causas. POLITO (1998)
expone que plagas, enfermedades, deficiencias nutricionales (Ca, B, Na), edad del
árbol, poda, estrés hídrico, incompatibilidad de polen o falta de polinización, no son
causales de una AFP en nogal.
Aunque es más severa en huertos de Serr, el AFP también se presenta en mayor o
menor escala en otros cultivares (POLITO, 1998).
Se puede esperar la AFP donde quiera que el cultivar Serr haya sido diseminado y
puede ocurrir con otros cultivares y plántulas en otras partes en menor grado y con
variaciones (CATLIN et al., 1987).
CATLIN et al. (1987) observaron que los pistilos de flores tomadas antes de la
detención del crecimiento en árboles que al final tuvieron 90% de AFP no observaron
daño celular, por lo que AFP no parece ser expresión de defectos inherentes del tejido
del ovario. El corte a través del ovario a pocos días de la detención del crecimiento no
reveló tejido necrótico. En las flores que abscisionaron se observó que cuando se
cortaron los ovarios de los pistilos para exponer los óvulos, éstos y el tejido placental
se presentaban necróticos, sugiriendo que el daño se originó dentro del pistilo y no
por causas externas (CATLIN y POLITO, 1989).
Los trabajos realizados desde 1990 por POLITO et al. (1998) muestran los siguientes
resultados:
- Tehama es ampliamente usado como cultivar polinizante en huertos Serr, sin
embargo, la incidencia del AFP siempre disminuye significativamente al
aumentar la distancia del polinizante. AFP generalmente alcanza un nivel estable
a 45 – 90 m desde el polinizante.
- El AFP es directamente proporcional a la carga de polen. Cuando se han
comparado los resultados de varios huertos en diferentes niveles de AFP, los

resultados revelan una relación consistente y altamente significativo entre al
número de granos de polen y el porcentaje de AFP.
- El AFP reduce el rendimiento, pero la producción se aumenta consistentemente
con la distancia desde el polinizante.
- Reduciendo la carga de polen se reduce el AFP y se incrementa la producción. Si
es posible disminuir a un 40% la AFP en un huerto, los rendimientos se verán
incrementados en más de un 60%.
2.4.2. Aborto de flores pistiladas en otros cultivares
Es sabido que la abscisión de flores pistiladas provocada por un exceso en la carga de
polen puede ocurrir, aunque en menor grado, en otros cultivares además de Serr
(CATLIN y OLSSON, 1990).
ROVIRA, NINOT y ALETA (2001) en estudios llevados a cabo en la colección de
germoplasma en IRTA-Mas Bové durante dos años, demostraron diferencias
significativas en la incidencia de AFP entre cultivares provenientes de cuatro
diferentes países: Estados Unidos, Chile, Francia y España. Así, en 19 cultivares
diferentes se llegó a la conclusión de que existen diferencias significativas entre años,
entre grupos de cultivares de diferente origen y dentro de los cultivares.
Al hacer una comparación de la AFP entre Serr, Sunland, Chandler y Howard,
durante siete años de estudio CATLIN y OLSSON (1990) encontraron que Serr tuvo
el porcentaje mayor de AFP, cercano al 80%, comparado con los otros tres cultivares
en donde se registraron valores del orden del 20% o menos, y no existió diferencia
entre ninguno de estos últimos.
En un estudio efectuado sobre cuatro cultivares de nogal Serr, Hartley, AS-1 y MB-T-
119, los resultados demostraron que la AFP en estos cuatro cultivares fue

significativamente diferente. El cultivar chileno AS-1 fue el menos afectado con un
2% de AFP y el cultivar español MB-T-119 tuvo la mayor proporción de AFP
cercana al 80%. En Hartley y Serr, de origen californiano, el porcentaje de AFP fue
cercano al 50% (ROVIRA y ALETA, 1997).
Según POLITO (1998) en un estudio encuentra que AFP en árboles de cv. Chandler y
cv. Vina adyacentes a los polinizadores fue de casi 40%. Chico presentó niveles de
AFP en árboles cercanos al polinizante de alrededor de 20 a 25%. De la información
anterior, POLITO et al. (1998) destacan que los cultivares Chandler, Vina y Chico
son menos sensibles que Serr a la dosis de polen, y cualquiera que sea la causa, la
reducida susceptibilidad a AFP tiene implicancias en las decisiones del manejo del
polen en el huerto. Señalan además, que las prácticas de disminución de polen que
son efectivas en un huerto de Serr, pueden tener sólo un efecto reducido en un huerto
de Chandler o de Vina.
2.4.3. Otras causas del aborto de flores y frutos en nogal
2.4.3.1 Peste negra del nogal
La bacteriosis del nogal (Juglans regia L.), enfermedad causada por Xanthomonas
campestris pv. Juglandis (Pierce), Dowson, más conocida por el nombre de “Peste
Negra” o “Walnut Blight”, constituye una de las principales limitantes de este cultivo
(ESTERIO, 1984).
Según PINILLA y ALVAREZ (1997), es la enfermedad más importante de este
cultivo en nuestro país, especialmente en aquellas regiones en donde las primaveras
se presentan lluviosas y templadas.

De acuerdo a lo anterior, la enfermedad es capaz de destruir un porcentaje
considerable de nueces, bajando en forma apreciable los rendimientos ( CARREÑO y
PINTO, 1988). Se ha visto que esta enfermedad ha recrudecido en forma catastrófica
llegando en algunos nocedales chilenos a pérdidas de hasta el 60% (ASTORGA y
ATORGA, 1987).
CARREÑO y PINTO (1988) determinaron que la bacteria es capaz de infectar fruta,
flores, brotes verdes, hojas y yemas. El ataque al fruto se puede producir muy
temprano en su desarrollo. En muchas ocasiones ocurre antes o durante la
polinización (LATORRE y ESTERIO, 1981).
Además, LATORRE y ESTERIO (1981) agregan que las pérdidas de mayor
consideración económica se producen como consecuencia de los ataques tempranos
debido a la caída prematura de flores pistiladas o de frutos recién cuajados.
PINILLA y ALVAREZ (1997) señalan que los síntomas consisten en el
atizonamiento y posterior ennegrecimiento de los tejidos tiernos o suculentos del
huésped, ya sea en amentos, flores pistiladas, brotes, foliolos, pecíolos, frutos, yemas
vegetativas y amentos durante su fase de latencia, como consecuencia del ataque se
ennegrecen parcial o totalmente.
Las condiciones favorables para que la bacteria se desarrolle es la presencia de agua
libre sobre los tejidos del huésped provocada por lluvias, lloviznas o neblinas
prolongadas, además de temperaturas que superen los 18°C. Por otra parte, cultivares
más precoces presentan una mayor suceptabilidad (PINILLA y ALVAREZ 1997).
Las medidas para controlar la peste negra están basadas fundamentalmente en la
prevención de la enfermedad, a través de prácticas culturales que incluyen, entre
otras, el control de malezas y la eliminación de frutos y amentos contaminados del
huerto (PINILLA y ALVAREZ 1997).

Además PINILLA y ALVAREZ (1997) afirman que agroquímicos con acción
bactericida, como los compuestos a base de cobre tal como oxicloruro de cobre, óxido
cuproso e hidróxido de cobre, deben ser aplicados para mantener protegidos los
tejidos tiernos durante los estados fenológicos de yema hinchada y elongación de
amentos. El número de pulverizaciones necesarias para mantener un buen control
varía de un año a otro. Se sugiere realizar por lo menos las tres primeras aplicaciones
destinas a proteger el árbol durante los primeros estados de desarrollo:
- prefloración, inmediatamente después de brotación;
- comienzo de la floración, justo al aparecer las primeras floraciones pistiladas,
pero antes de plena flor, y
- postfloración, una vez que las flores pistiladas estén polinizadas.
2.4.3.2. Polilla de la manzana
Cydia pomonella, conocida como polilla de la nuez o de la manzana, según ESTAY
(1997), es la más importante en nuestro país por su carácter primaria y cuarentenaria,
además de producir un daño directo en los frutos.
De acuerdo a PRADO (1998) con respecto a la fenología del insecto, éste inverna
como larva de último estadío. A comienzos de octubre comienzan a nacer los
primeros ejemplares machos, teniendo un máximo de capturas en trampas la tercera
semana de octubre. Ocho a diez días después de la aparición de los machos nacen las
hembras las cuales se aparean dos días después de emergidas, siempre que las
temperaturas sean superiores a 15°C. La ovipostura en general se extiende hasta
mediados de febrero por el traslapo de generaciones (ESTAY, 1997).
La polilla presenta tres generaciones en Chile. La primera generación llega a un
máximo de captura a mediados de diciembre. La segunda generación produce un
máximo de vuelo a mediados de enero y dañará los frutos a partir de fines de enero.
Una tercera generación, se produce en febrero y parte de marzo dañando frutos

durante febrero y marzo. La primera y, especialmente, la segunda penetración de
frutos son las más dañinas (PRADO, 1998).
Según ESTAY (1997) la primera generación reduce directamente los rendimientos,
porque causa la caída de frutos recién cuajados. Este daño de la polilla podría
confundirse con la caída natural de frutos (no fertilizados), o con el problema de peste
negra, los que presentan lesiones necróticas y que a pesar de producirse en la misma
época se distingue por los residuos dejados por la plaga. La segunda generación
afecta directamente la calidad de la nuez y el rendimiento comercial.
Para un correcto manejo, PRADO (1998) establece que el huerto debe monitorearse
con trampas para la polilla de la manzana.
La trampa con feromona permite conocer el inicio y el término del vuelo y el nivel de
infestación. Seis trampas, independiente del número de hectáreas, debe dar una buena
estimación del vuelo de la polilla. Deben ser ubicadas siempre en el interior del
huerto a un mínimo de 150 metros entre ellas. Se deben colocar la segunda quincena
de septiembre utilizando Codlemone como feromona, ésta debe ser cambiada cada
seis semanas y la base de las trampas tres veces durante la temporada. Las trampas
deben ser revisadas dos veces por semana, retirando las polillas, y llevando un
recuento para determinar el período de mayor caída diaria de machos por trampa
(ESTAY, 1997).
De acuerdo a lo anterior, PRADO (1998) señala que los insecticidas deben aplicarse
en el lapso de tiempo entre la eclosión de la larva y la penetración del fruto,
correspondiendo a 8 - 10 días después del máximo de captura. Los insecticidas más
usados corresponden a Azinphos methyl, Phosmet, Clorpirifos, Metidathion y
Diazinon.

3. MATERIALES Y MÉTODOS
3.1. Ubicación del ensayo:
El estudio fue realizado entre septiembre de 2002 y febrero de 2003, en el fundo “Los
Tilos”, ubicado en la comuna de Hijuelas, Provincia de Quillota, V Región, Chile. La
localización geográfica corresponde a 32°45’ latitud Sur y 71°11’ longitud Oeste y
se encuentra ubicado a 253 m.s.n.m.
3.2. Caracterización de la zona del ensayo:
3.2.1. Clima
La localidad de Hijuelas cuenta con un clima de tipo templado Mediterráneo
semiárido. El régimen térmico de esta localidad se caracteriza por temperaturas que
varían, en promedio, entre una máxima en enero de 27,7ºC y una mínima de julio de
4,7ºC. El periodo libre de heladas es de 245 días, con un promedio de siete heladas
por año. Registra anualmente 1650 días-grado y 977 horas de frío. El régimen hídrico
se caracteriza por una precipitación media anual de 454 mm y un periodo seco de
ocho meses. Esta localidad presenta influencia oceánica, por lo que las condiciones
climáticas térmicas se ven atenuadas (SANTIBAÑEZ y URIBE, 1990).
3.2.2. Suelo
El suelo de la localidad de Hijuelas es sedimentario, de origen coluvial, derivado de
rocas neutras a básicas, profundo, de textura franca y de color pardo a pardo oscuro
en la superficie; textura franca, de color pardo a pardo oscuro en profundidad.
Sustrato constituido por gravas, gravillas y bolones con matriz franco arcillo arenosa.

Suelo de topografía suavemente inclinada (piedmont), de permeabilidad moderada y
de buen drenaje (CIREN-CORFO, 1998).
3.3. Material vegetal:
El estudio se llevó a cabo en 24 árboles de nogal, cultivar Serr, de una plantación
realizada en 1994 e injertados sobre patrón Juglans regia, con un marco de plantación
definitivo de 8,5 x 8,5 m en hileras intercalas con árboles de cultivar Chandler. Los
árboles fueron conducidos con un sistema de eje central.
Al momento de la selección de los árboles en la ejecución de los distintos
tratamientos, se eligieron aquellos árboles de apariencia saludable y representativos
de la condición del huerto.
Los árboles son regados con un sistema de microaspersión, con un caudal de 56
Lhora-1, cada árbol es regado por dos emisores. Los emisores están ubicados a una
distancia de 1 – 1.50 m del tronco del árbol.
Durante la temporada 2002 en el huerto se efectuaron labores culturales propias del
manejo, las que se pueden observar en el Anexo 2.
3.4. Ensayos realizados :
3.4.1. Determinación de la fenología floral del cv. Serr:
Se realizó un seguimiento de la fenología de la floración, tanto de flores femeninas y
masculinas, siguiendo los estados descritos por MUNCHARAZ (2001).
Se seleccionaron ocho árboles al azar, en cada árbol fueron marcados 10 amentos y
10 brotes con flores femeninas. Las distintas estructuras fueron seleccionadas al azar

y comprendían todo el perímetro del árbol tanto dentro y fuera de éste. Para llevar el
registro fenológico floral, se consideró el árbol en su conjunto mediante apreciación
visual. Los estados fenológicos identificados son los siguientes (MUNCHARAZ,
2001):
• Para las inflorescencias masculinas:
1. Inicio de liberación de polen: Total amarillamiento de las anteras y comienzo de
la dehiscencia de éstas, desde la base del amento (Fm).
2. Plena liberación de polen (más del 50% de los amentos se encuentran liberando
polen): Dehiscencia total de las anteras y plena emisión de polen (Fm2).
3. Término de liberación de polen: Las anteras, ya sin polen, toman color pardo
(Gm).
• Para las flores femeninas:
1. Inicio de receptividad del estigma: Aparecen los estigmas, que comienzan a
separarse y pasan de rojo a verde – amarillento. Comienza su receptividad, que en
este caso es máxima (Ff1).
2. Plena receptividad del estigma (más del 50% de las flores femeninas se
encuentran receptivas): Los estigmas se abren y finalizan su receptividad. Su
color es verde pálido (Ff2).
3. Término de receptividad del estigma: Marchitamiento de los estigmas. Se
desecan, pasando del marrón al negro. Ya no son receptivos (Gf)
(MUNCHARAZ, 2001).
Se consideró como peak de la floración masculina y femenina cuando más del 50%
de los árboles estudiados se encontraran en plena liberación de polen y plena
receptividad del estigma, respectivamente.

3.4.2. Determinación de la efectividad del tratamiento de agitación sobre la caída de amentos y carga de polen:
Se trabajó con cuatro árboles por tratamiento y con dos bloques. Dentro de cada uno
de éstos se encontraban el tratamiento de agitación más el tratamiento testigo. Los
tratamientos realizados se describen a continuación:
T0: Tratamiento testigo
T1: Se realizó la agitación con las primeras flores femeninas receptivas (menor o
igual al 5%).
Se consideró remecer todo el sector seleccionado para cada uno de los tratamientos,
para evitar la contaminación con polen de los otros árboles. La superficie remecida,
contabilizando ambos bloques fue de 4.2 ha con un total de 454 plantas.
Para determinar la intensidad de agitación, se realizaron pruebas en árboles fuera del
sector en estudio. Se identificaron 10 ramillas y se contabilizó el total de amentos.
Para ésto se consideró un porcentaje de caída de amentos de aproximadamente 50%.
Este porcentaje estaría de acuerdo a los establecido por KRUEGER (2001) donde
propone remover hasta un 80% de los amentos, que corresponde al momento en que
los primeros amentos han caído del árbol .
Para realizar la agitación propiamente tal se utilizó una remecedora acoplada a los
tres puntos del tractor que es accionada por el eje toma fuerza de éste. La faja se
ubicó alrededor del tronco a una altura de 80 cm aproximadamente desde el nivel del
suelo. El tiempo de remecido fue de 1 segundo por árbol, con una intensidad de
remecido de 1000 rpm.
La carga de polen se midió indirectamente mediante el conteo de amentos. Esta labor
se hizo tanto en árboles testigo como en los del tratamiento. La medición se hizo cada

tres días contando la totalidad de amentos caídos al suelo. Para los árboles
correspondientes al tratamiento, los amentos se contabilizaron antes, inmediatamente
después y durante las mediciones posteriores.
Con el propósito de agilizar el conteo de amentos, se colocó una lona plástica
provisoria sólo bajo los árboles propios del tratamiento y sólo al minuto de agitación
de los árboles, con el fin de recoger más rápido los amentos y colocarlos en un
cartucho de papel previamente identificado.
3.4.3. Determinación del aborto de flores pistiladas :
Las variables de respuesta registradas durante esta temporada, influídas por el
tratamiento de agitación aplicado a los árboles de nogal, cultivar Serr fueron:
• Aborto de flores pistiladas con diámetro de 3 a 4 mm
Cuando el ovario presentó un diámetro entre 1,5 mm y 2 mm comenzaron las
mediciones al ovario de las flores etiquetadas (25 de septiembre, 2002), debido a que
el cese del crecimiento del ovario es a los 3 ó 4 mm produciendo un aborto en las
flores pistiladas (CATLIN y POLITO, 1989). De acuerdo a esto, la cuantificación de
las mediciones se basó en los diámetros anteriormente señalados.
• Aborto de flores pistiladas con diámetro mayor o igual a 7mm
La medición del ovario de las flores comenzó cuando éstos tenían un diámetro de 5
milímetros (8 octubre, 2002), hasta un diámetro de 30 mm a mediados de noviembre
del mismo año. KRUEGER (2000) especifica que la pérdida de flores pistiladas

ocurre dos a tres semanas después de la pérdida por exceso de polen y con un
diámetro de ovario mayor o igual a 7 mm.
• Aborto de flores pistiladas por otras causas
Dentro de esta categoría están aquellas flores y/o frutos afectados por polilla de la
manzana y peste negra del nogal. Cydia pomonella produce daños directos a los
frutos y causa la caída de frutos recién cuajados. Un signo de la presencia de la plaga
son los residuos dejados por ésta en la fruta. Xanthomonas campestris pv. Juglandis
infecta fruto, flores, brotes y yemas, causando caída prematura de frutos recién
cuajados. Los síntomas consisten en el atizonamiento y posterior ennegrecimiento de
los tejidos.
Las mediciones del ovario se efectuaron desde mediados de noviembre del 2002 hasta
mediados de febrero del 2003.
• Frutos retenidos en el árbol
Esta medición se realizó a mediados de febrero del 2003.
Para la determinación del aborto de flores pistiladas femeninas, se estimó los
porcentajes (mediante el promedio), de cada árbol a partir de la información obtenida
de 10 brotes seleccionados al azar, con 1-5 flores femeninas por brote, por lo tanto, se
trabajó con 15 a 30 flores femeninas por unidad experimental (árbol). Los brotes
fueron etiquetados a principios de brotación, cuando las primeras hojas estaban
completamente abiertas y erguidas, toman un porte oblicuo y dejan aparecer las flores
femeninas (Df2).

Se llevó un registro del número de flores femeninas por brote y tamaño de los
ovarios, tanto para la variable aborto de flores con diámetro entre 3-4 mm como para
la variable aborto de flores con diámetro mayor o igual a 7mm. La medición se hizo
cada tres a cuatro días, registrando el diámetro de éstos, utilizando un pie de metro.
3.4.4. Diseño experimental:
El fundo de Hijuelas tiene una pendiente de un 4%, lo que justifica dividirlo en dos
partes (bloques). Dentro de cada bloque se seleccionó al azar los sectores del
tratamiento y los árboles propios del tratamiento. La experiencia se realizó mediante
un diseño en bloques completamente al azar.
La Figura 3 muestra la ubicación del tratamiento de agitación y el de los árboles
testigos dentro del huerto de nogales cv Serr, Hijuelas, V región.

Viento N
Tratamiento 1
Bloque 2
(4 árboles)
Testigo 2
(4 árboles)
Tratamiento 1
Bloque 1
(4 árboles)
Testigo 1
(4 árboles)
FIGURA 3. Esquema de la ubicación de los tratamientos de agitación en árboles de nogal cv. Serr en el huerto ubicado en Hijuelas.

3.4.4.1. Análisis de la información
Para la realización del análisis de varianza se comprobó el cumplimiento del supuesto
de normalidad de las observaciones, lo cual se realizó mediante la prueba de
Kolmogorov-Smirnov. De no comprobarse la normalidad de los datos se recurrió a la
prueba no paramétrica de Kruskal – Wallis. Se trabajó con un nivel de significación
del 5%.
Para cada variable de respuesta el modelo propuesto es el siguiente:
Yijk = µ + Ti + Bj + εijk
Con i = 1,3; j = 1,2; k = 1,3; donde Yijk es el k-ésimo porcentaje de la variable de
respuesta con la aplicación del tratamiento (i) en el bloque (j), µ es la media general,
Ti es el efecto causado por el i-ésimo “tratamiento de agitación”, Bj es el efecto
causado por el j-ésimo “bloque”, εijk es el k-ésimo error experimental aleatorio en el
tratamiento (i) en el bloque (j). Para cada variable de interés la hipótesis es la
siguiente:
H0: Ti = 0 = No existe efecto del tratamiento de agitación sobre la variable de
respuesta correspondiente en nogales cv. Serr.
En caso de existir efecto de los tratamientos sobre algunas de las variables de
respuesta, se procedió a realizar una separación de medias mediante del test de
Duncan con α = 0.05.
La realización del análisis estadístico se realizó por medio del Software Estadístico
SPSS versión 6.1 y de la planilla de cálculo MS EXCEL 97 (Microsoft).

3.5. Análisis de registros climáticos:
El seguimiento fenológico descrito se relacionó con las variables climáticas más
influyentes sobre la fenología floral: temperatura, humedad relativa y precipitación.
También se registró la velocidad del viento y su dirección. Se utilizó el registro diario
de estas variables a través de los registros de la estación metereológica ‘Los Tilos’
(LTS) ubicada en el interior del huerto del mismo nombre.

4. PRESENTACIÓN Y DISCUSIÓN DE LOS RESULTADOS
4.1. Fenología floral del nogal cv. Serr:
La floración masculina comenzó el día 4 de septiembre de 2002 y se extendió hasta el
30 de septiembre del mismo año. La flor femenina comenzó su receptividad el día 22
de septiembre extendiéndose hasta el 4 de octubre del 2002. En el Figura 2, que
muestra la fenología de la floración femenina y masculina de nogales cv. Serr, se
puede observar que se produjo un período de traslape entre la floración masculina y
femenina de nueve días durante la temporada 2002. La figura 4 muestra la totalidad
de la floración tanto femenina como masculina, sin considerar períodos peak de
liberación de polen o peak de receptividad del estigma. Es posible comparar de esta
manera el traslape de floración, éste muestra que durante el año 2001 la floración
femenina estuvo cubierta durante 30 días por la floración masculina, al contrario de
este año, que sólo estuvo cubierta por nueve días.
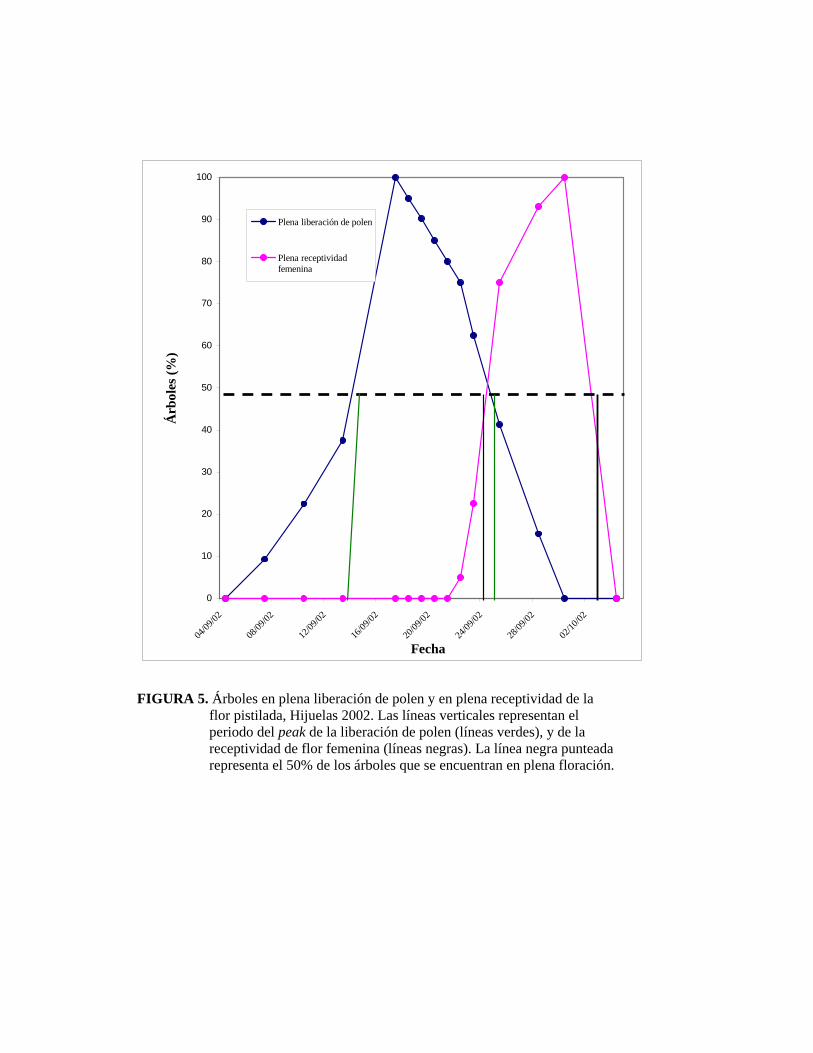
La Figura 5 muestra el registro de árboles en plena liberación de polen y plena
receptividad de la flor femenina, considerando como pleno aquel estado en donde
más del 50% de los amentos se encuentran liberando polen y más del 50% de las
flores femeninas se encuentran receptivas. Se puede observar que se produjo un
periodo de traslape entre la plena floración masculina y la plena floración femenina
de 3 días. Si bien este período es muy corto, hay que destacar que en este caso se está
en el pleno de floración masculina y femenina, es decir, cuando más del 50% de los
árboles estudiados están en plena liberación de polen y plena receptividad del
estigma, respectivamente; y no tomando la totalidad de la floración femenina y
masculina, ya que si fuera asi, el período y porcentaje de cubrimiento de la floración
femenina sería mayor.

Al comparar los registros obtenidos por HERRERA (2001), se observa que en la
presente temporada la floración ocurrió 16 días antes en relación al año pasado en el
mismo huerto (Anexo 3).

FIGURA 4. Fenología de la floración femenina y masculina de nogales cv. Serr. Los datos fueron recolectados en el año 2002 en la localidad de Hijuelas, V Región.
Hijuelas Septiembre Octubre
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 1 2 3 4
Liberaci
ón de
Polen
X X X X X X X X X X X X X X X X X X X X X X X X X X X
Receptiv
idad
Estigma
X X X
X X X X X X X X XX
0
10
20
30
40
50
60
70
80
90
100
04/09
/02
08/09
/02
12/09
/02
16/09
/02
20/09
/02
24/09
/02
28/09
/02
02/10
/02
Fecha
Árb
oles
(%)
Plena liberación de polen
Plena receptividadfemenina
FIGURA 5. Árboles en plena liberación de polen y en plena receptividad de la flor pistilada, Hijuelas 2002. Las líneas verticales representan el periodo del peak de la liberación de polen (líneas verdes), y de la receptividad de flor femenina (líneas negras). La línea negra punteada representa el 50% de los árboles que se encuentran en plena floración.

Otras publicaciones han constatado el hecho de tener distintos comportamientos
de floración en cuanto a su extensión y traslape en otras zonas del país,
VALENZUELA, LOBATO y LEMUS (2001) y ARAYA (2000).
De la misma manera, en un estudio de biología floral en distintas variedades
realizado durante cinco años en la Universidad de California, reveló que no se
produce un traslape completo entre los periodos de floración masculina y
femenina, siendo en algunas variedades de sólo cinco días y sin coincidencia de
los peaks de floración (HENDRICKS et al., 1998).
ARAYA (2000), reportó un traslape de floración femenina y masculina de un
100% en la zona de Talagante, debido probablemente a una baja acumulación
térmica presente en la zona, lo que habría extendido la floración masculina.
POLITO (1998) expone que la abscisión de flores pistiladas varía mucho de
cultivar a cultivar, de lugar a lugar y de año a año. Es por esta razón que un
productor necesita ser capaz de predecir la extensión del traslape de la floración
para determinar la factibilidad de remover los amentos.
4.2. Efectividad del tratamiento de agitación sobre la caída de amentos y
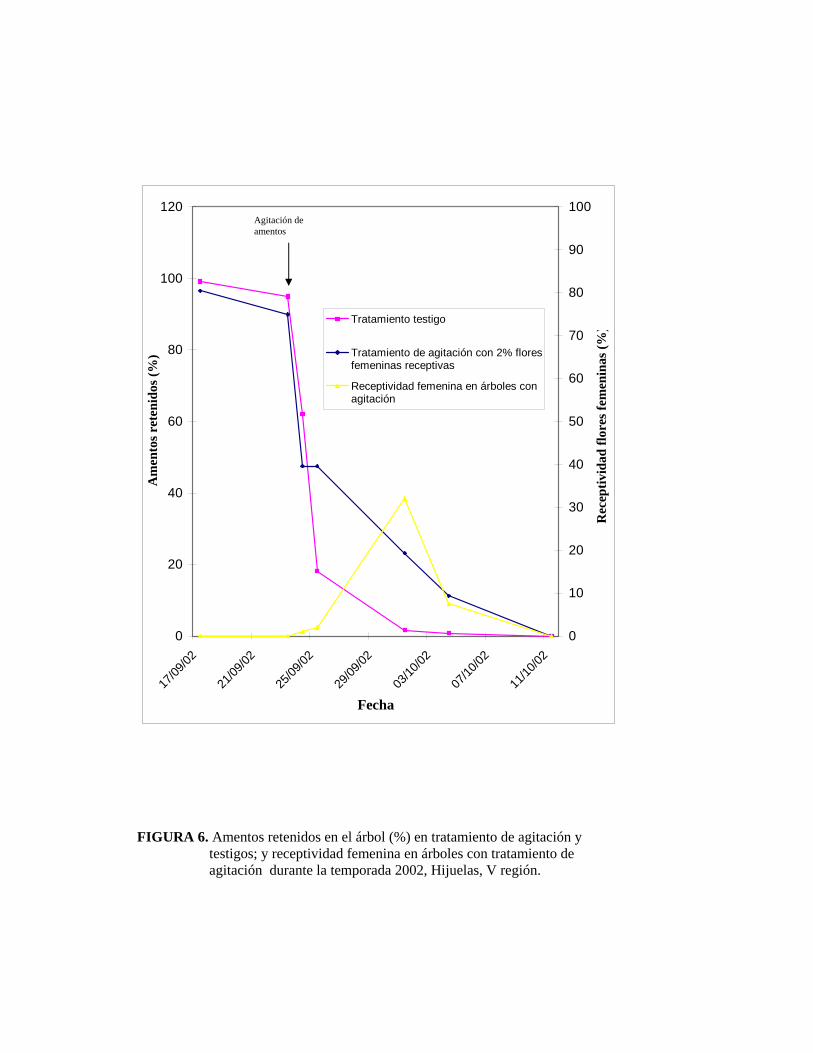
carga de polen:
El tratamiento de agitación se efectúo cuando había caído un 37 % de amentos y
existía un 2,3 % de flores femeninas receptivas en los árboles del tratamiento.
Aún cuando al momento de remoción de amentos, el 63% de éstos se encontraba
en el árbol, gran parte de ellos estaba en el estado de término de liberación de
polen; por lo tanto, la carga de polen que recibió el 2% de flores femeninas fue
baja.
La Figura 6 muestra los amentos retenidos en el árbol (%) durante la temporada
2002 en Hijuelas.
0
20
40
60
80
100
120
17/09
/02
21/09
/02
25/09
/02
29/09
/02
03/10
/02
07/10
/02
11/10
/02
Fecha
Am
ento
s ret
enid
os (%
)
0
10
20
30
40
50
60
70
80
90
100
Rec
eptiv
idad
flor
es fe
men
inas
(%)
Tratamiento testigo
Tratamiento de agitación con 2% floresfemeninas receptivas
Receptividad femenina en árboles conagitación
FIGURA 6. Amentos retenidos en el árbol (%) en tratamiento de agitación y testigos; y receptividad femenina en árboles con tratamiento de agitación durante la temporada 2002, Hijuelas, V región.
Agitación de amentos

Los amentos comenzaron a caer desde el 17 de septiembre extendiéndose hasta el
11 de octubre, tanto en árboles testigos como en árboles donde se aplicó el
tratamiento de agitación.
La diferencia entre el comportamiento de estas dos curvas (tratamiento de
agitación versus testigo), radica principalmente en el tiempo de permanencia de
amentos liberando polen en el árbol y abundancia de éstos, en donde los árboles
testigos demostraron un mayor tiempo con amentos retenidos.
Al minuto de hacer la remoción de amentos, la receptividad femenina estaba
recién comenzando, y la cantidad de amentos que quedaron en el árbol (20%) fue
suficiente para polinizar y fecundar al resto de las flores femeninas receptivas, lo
cual se corrobora con lo concluído por KRUEGER (2000), en donde describe que
la fecundación se asegura hasta con un 20% de amentos en los árboles.
Un 32% de las flores femeninas presentaba receptividad el día 30 de septiembre,
cuando quedaba menos de un 5% de amentos retenidos en el árbol.
Esta situación es distinta a la ocurrida el año anterior, en donde los tratamientos se
efectuaron con un 15% y un 50% de flores femeninas receptivas, lo cual era un
porcentaje elevado de receptividad de flores pistiladas. Este porcentaje de
receptividad femenina es elevado, debido a que la gran mayoría de las flores
femeninas ya fueron polinizadas con un exceso de polen, lo cual trae como
consecuencia un mayor aborto de flores femeninas por esta causa.
POLITO (1998) expone que el tratamiento de remoción se puede efectuar en dos
situaciones: teniendo una coincidencia marcada (en período y magnitud) de
traslape de floración femenina y masculina o, en el caso contrario, en donde existe
baja coincidencia de floración en el cultivar Serr. En estos dos casos la remoción
de amentos ayuda a bajar las pérdidas de aborto de flores pistiladas por exceso de
polen.

Al comparar la carga de amentos en árboles testigos, entre la temporada 2001 y
2002, es posible determinar que durante la presente temporada se registró un 38%
más que lo registrado en la temporada anterior. Durante la temporada 2001 se
registró un total de amentos por planta de 3530 unidades, mientras que durante la
última temporada fue de 5744.
Es posible remover la mayor parte de los amentos cuando los primeros han caído,
ya que en ese momento ha comenzado la descarga de polen, pero la mayoría no
han liberado su polen y pueden ser removidos sin demasiado esfuerzo
(KRUEGER, 2000). Sin embargo, LEMUS, VALENZUELA y LOBATO (2001)
recomiendan remecer los árboles cuando aparece la primera flor femenina,
considerando el peak de liberación de polen en ese momento. Al momento de
hacer el tratamiento existía un bajo porcentaje de flores receptivas durante la
plena descarga de polen, por lo que se puede considerar como una fecha precisa
para la remoción de amentos. (Figura 7).
4.3. Aborto de flores pistiladas:
4.3.1. Aborto de flores pistiladas con diámetro entre 3 a 4 milímetros
POLITO (1998) afirma que el número de flores abortadas por brote, es
independiente al número total de éstas por brote.
Durante la última semana de septiembre fue posible observar las primeras flores
femeninas receptivas con un diámetro de ovario cercano a 3 mm. Las primeras
flores abortadas se observaron el 11 de octubre (Figuras 8 y 9), concordando de
esta manera con CATLIN y OLSSON (1990) quienes señalan que la abscisión por
exceso de polen ocurre una a dos semanas después de la detención del crecimiento
del ovario, a los 3 ó 4 mm de diámetro; que en este estudio ocurrió dos semanas
después de ser polinizadas.

La Figura 9 muestra la diferencia de diámetros que existe entre un fruto
polinizado con exceso de polen y un fruto polinizado con falta de polen.
El Cuadro 1 muestra valores de aborto de flores pistiladas con diámetro 3-4
milímetros, los valores son expresados en porcentaje.
CUADRO 1. Efectividad del tratamiento de agitación del tronco para reducir la carga de polen, sobre el aborto de flores pistiladas en nogal cv. Serr. Hijuelas, 2002.
TRATAMIENTO ABORTO DE FLORES PISTILADAS
CON DIÁMETRO ENTRE 3-4 mm (%)
Testigo 24,01 b 1
Agitación 9,21 a
1 Valores seguidos de una misma letra no difieren estadísticamente según test de Duncan
(α= 0.05).
El bajo porcentaje de aborto de flores pistiladas por un exceso de polen en los
árboles testigos se debió principalmente al corto traslape entre la floración
masculina y femenina que existió la temporada 2002. El tratamiento de remoción
fue efectivo, ya que se logró disminuir el polen disponible al momento en que las
flores femeninas estuvieron receptivas, lo que redujo la caída de frutos recién
cuajados.

FIGURA 7. Estado de desarrollo de los árboles al momento de realizar el
tratamiento de agitación (25 septiembre del 2002). Bajo el árbol se instaló una película de polietileno para recibir los amentos luego de la agitación. La línea negra indica la posición de un cable de acero del agitador mecánico unido al tractor que produce la vibración del tronco y la posterior caída de amentos.
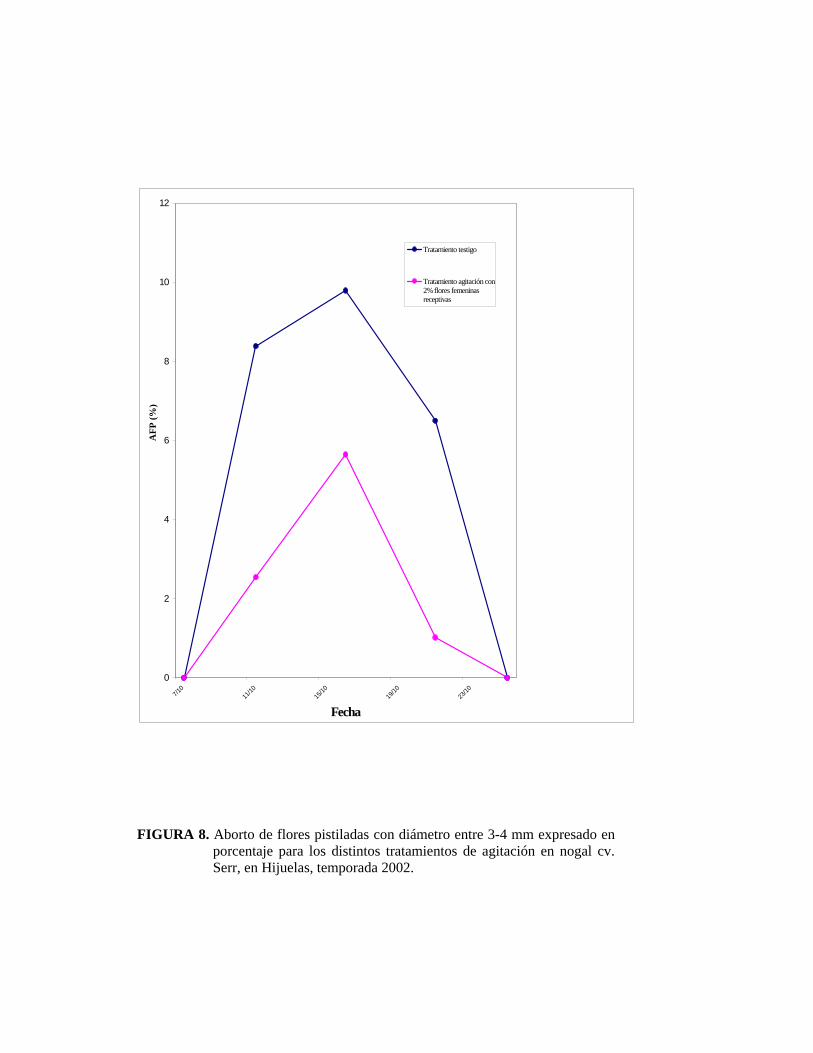
FIGURA 8. Aborto de flores pistiladas con diámetro entre 3-4 mm expresado en
porcentaje para los distintos tratamientos de agitación en nogal cv. Serr, en Hijuelas, temporada 2002.
0
2
4
6
8
10
12
7/10
11/10
15/10
19/10
23/10
Fecha
AFP
(%)
Tratamiento testigo
Tratamiento agitación con2% flores femeninasreceptivas

FIGURA 9. A. Fruto de nogal abortado por un exceso de polen con diámetro de 3
mm (a la izquierda del brote). B. Diferencia de diámetros entre frutos abortados por exceso de polen (a la izquierda del brote, diámetro 3mm) y por falta de polen (a la derecha del brote, diámetro 7 mm).
A B

En el presente ensayo el tratamiento de agitación permitió reducir la caída de
frutos recién cuajados al 9,2%, en comparación con el testigo en que se obtuvo un
24%. Estos resultados son diferentes a los obtenidos por HERRERA (2001), quien
registra valores entre 50% y 43% de aborto de frutos recién cuajados en los
tratamientos de agitación realizados con 15% y 50% de flores femeninas
receptivas.
Los árboles testigo presentaron un 70% de aborto de flores pistiladas; sumándose
así a resultados obtenidos por CATLIN Y OLSSON (1990) en donde se observan
valores expresados en porcentajes por exceso de polen entre un 27% y 80%. Del
mismo modo, McGRANAHAN et al. (1994) publicaron resultados de AFP en el
año 1990 en el cultivar Serr de 93%, en el año 1991 de 33% y en el año 1992 de
86%. Por su parte ROVIRA y ALETA (1997) registraron AFP del orden del 60%
al 90%.
4.3.2. Aborto de flores con diámetro mayor o igual a 7 milímetros
Las flores que no fueron polinizadas continuaron su crecimiento cerca de tres
semanas antes que cayeran del árbol. Éstas presentaban un diámetro de ovario
mayor que 3 mm y el pedúnculo quedaba unido al ápice del brote (POLITO y LI,
1985; CATLIN et al., 1987), además los estigmas mantuvieron su color y mucho
de su forma original (POLITO 1998).
Durante la segunda y tercera semana de octubre se concentraron las caídas de
fruto con diámetro igual o superior a 7 milímetros (Figura 10), coincidiendo a la
vez con las caídas por exceso de polen. La Figura 11 muestra la diferencia que
existe en diámetro de fruto y adherencia de pedúnculo, entre frutos abortados por
exceso de polen (diámetro 3 mm) y por falta de polen (diámetro mayor o igual a
7 mm).

Si bien las caídas por falta de polen se tienden a concentrar en un lapso de tiempo
determinado, no es fácil distinguir una fecha con precisión.
En la presente temporada las caídas tuvieron dos peaks, la segunda y cuarta
semana de septiembre. Esto confirma lo expresado por POLITO y LI (1985)
quienes afirman que las flores que abortan por una falta de polen pueden quedar
adheridas al árbol durante tres semanas o más.
El Cuadro 2 señala el aborto de flores pistiladas con un diámetro igual o superior
a 7 milímetros, en árboles testigos y en árboles con tratamiento de agitación
CUADRO 2. Efectividad del tratamiento de agitación del tronco para reducir la carga de polen, sobre el aborto de flores pistiladas en nogal cv.
Serr. Hijuelas, 2002.
TRATAMIENTO ABORTO DE FLORES (%) CON
DIÁMETRO MAYOR O IGUAL 7 mm
Testigo 3,55 NS
Agitación 3,17
NS: no significativo (p>0.05 en el análisis de varianza)
Del cuadro anterior se puede inferir que no existe diferencia entre la absición de
flores pistiladas de testigos y el tratamiento. Este resultado difiere con lo descrito
por HERRERA (2002) durante la temporada 2001, en donde se observan valores
para los testigos de 9% y para los tratamientos un 6,33% en promedio; si bien la
diferencia tampoco fue significativa estadísticamente, sí existe diferencia en
valores registrados entre una temporada y otra.

FIGURA 10. Aborto con diámetro mayor o igual a 7 mm, para los distintos tratamientos de agitación en nogal cv. Serr, en Hijuelas, durante la temporada 2002.
0
0,2
0,4
0,6
0,8
1
1,2
1,4
1,6
1,8
7-10
10-10
13-10
16-10
19-10
22-10
25-10
28-10
31-10 3-1
16-1
19-1
112
-1115
-11
Fecha
AFP
por
falta
de
pole
n (%
)
Tratamiento testigo
Tratamiento de agitación con 2% deflores femeninas receptivas

FIGURA 11. Frutos de nogal abortados por exceso y falta de polen. A.
Comparación entre varios frutos abortados. Además se diferencia la adherencia de pedúnculo en un fruto abortado por exceso de polen versus uno abortado por falta de polen, éste no presenta pedúnculo adherido. B. Detalle de un par de frutos abortados por dos causas distintas, exceso de polen (3 mm) y falta de polen (mayor o igual 7 mm). La flecha en A muestra un fruto abortado por exceso de polen con diámetro de 3 mm. La flecha en B indica la ausencia de pedúnculo en un fruto abortado por falta de polen Cada cuadro representa 1 mm2.
A
B

4.3.3. Aborto de flores por otras causas
El promedio obtenido entre ambos tratamientos es alto (Cuadro 3), lo que indica
que el manejo contra plagas y enfermedades del cultivo no fue lo suficiente para
aminorar estas pérdidas. El alto porcentaje de fruta dañada corresponde a daño por
polilla y peste negra, por lo que es una causa importante de aborto de frutos y en
la reducción de la productividad en el cultivar. Estos frutos caían con diámetros
mayores a 35 mm, por lo que queda descartada la posibilidad que sean frutos
abortados por un exceso de polen (CATLIN et al., 1987).
CUADRO 3. Aborto de flores pistiladas de nogal cv. Serr por otras causas en Hijuelas. (%)
TRATAMIENTO ABORTO DE FLORES (%) POR OTRAS CAUSAS
Testigo 1,63 NS
Agitación 3,70
NS: no significativo (p> 0.05 en el análisis de varianza).
En la Figura 12 se puede observar gráficamente la caída de flores por otras causas,
viendo una notable concentración a mediados de febrero. Esta caída esta
influenciada básicamente por la segunda y tercera generación de polilla de la
manzana, donde son dañados los frutos desde fines de enero hasta inicios de
marzo.
En la Figura 13 se observa un resumen de las diversas caídas registradas en la
temporada 2002. Estos datos se pueden comparar con los datos que se muestran
en el Anexo 6, que corresponde a un resumen de las diversas caídas de flores
pistiladas durante la temporada 2001. El comportamiento de las variables aborto
de flores con diámetro entre 3 – 4 mm y aborto de flores con diámetro mayor o
igual a 7 mm ( en los árboles testigo), fue similar en ambas temporadas
(2001/2002).

No obstante las caídas esta temporada 2002 ocurrieron un poco antes debido a que
la floración ocurrió antes que la temporada 2001.
4.3.4. Retención de frutos
En el Cuadro 4 se puede visualizar que existe una diferencia significativa en el
número de frutos que quedaron en el árbol entre árboles testigos y tratamientos,
luego de la temporada de estudio 2002.
En comparación con los datos obtenidos por HERRERA (2002) en la presente
temporada hubo mayor retención de frutos, debido principalmente a la mayor
respuesta del tratamiento de remoción.
CUADRO 4: Frutos retenidos hasta la cosecha en Hijuelas.
TRATAMIENTO FRUTOS RETENIDOS EN EL ÁRBOL (%)
Testigo 70,80 a 1
Agitación 83,91 b 1 Valores seguidos de una misma letra no difieren estadísticamente según test de Duncan (α = 0.05).
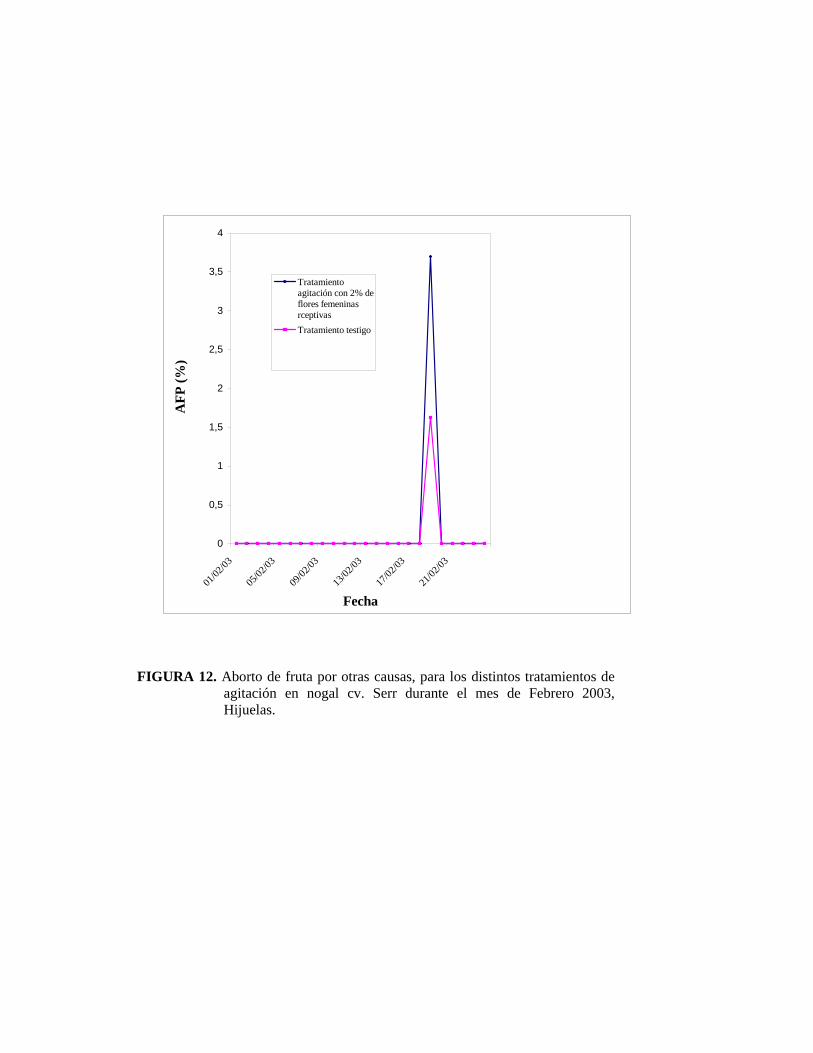
FIGURA 12. Aborto de fruta por otras causas, para los distintos tratamientos de agitación en nogal cv. Serr durante el mes de Febrero 2003, Hijuelas.
0
0,5
1
1,5
2
2,5
3
3,5
4
01/02
/03
05/02
/03
09/02
/03
13/02
/03
17/02
/03
21/02
/03
Fecha
AFP
(%)
Tratamientoagitación con 2% deflores femeninasrceptivasTratamiento testigo

0
0,5
1
1,5
2
2,5
3
3,5
4
4,5
5
5,5
6
6,5
7
7,5
8
8,5
9
9,5
10
10,5
07/10
/02
17/10
/02
27/10
/02
06/11
/02
16/11
/02
26/11
/02
06/12
/02
16/12
/02
26/12
/02
05/01
/03
15/01
/03
25/01
/03
04/02
/03
14/02
/03
24/02
/03
Fecha
Caí
das d
e flo
res p
istil
adas
(%)
Aborto de flores pistiladas con diámetro 3-4 mm"
Aborto de flores pistiladas con diámetro mayor o igual a7 mm
Aborto de flores pistiladas por otras causas
FIGURA 13. Porcentaje de aborto de flores por diversas causas en huerto de nogal cv. Serr ubicado en la localidad de Hijuelas, V Región, durante la temporada 2002/2003, sin aplicación de tratamientos.
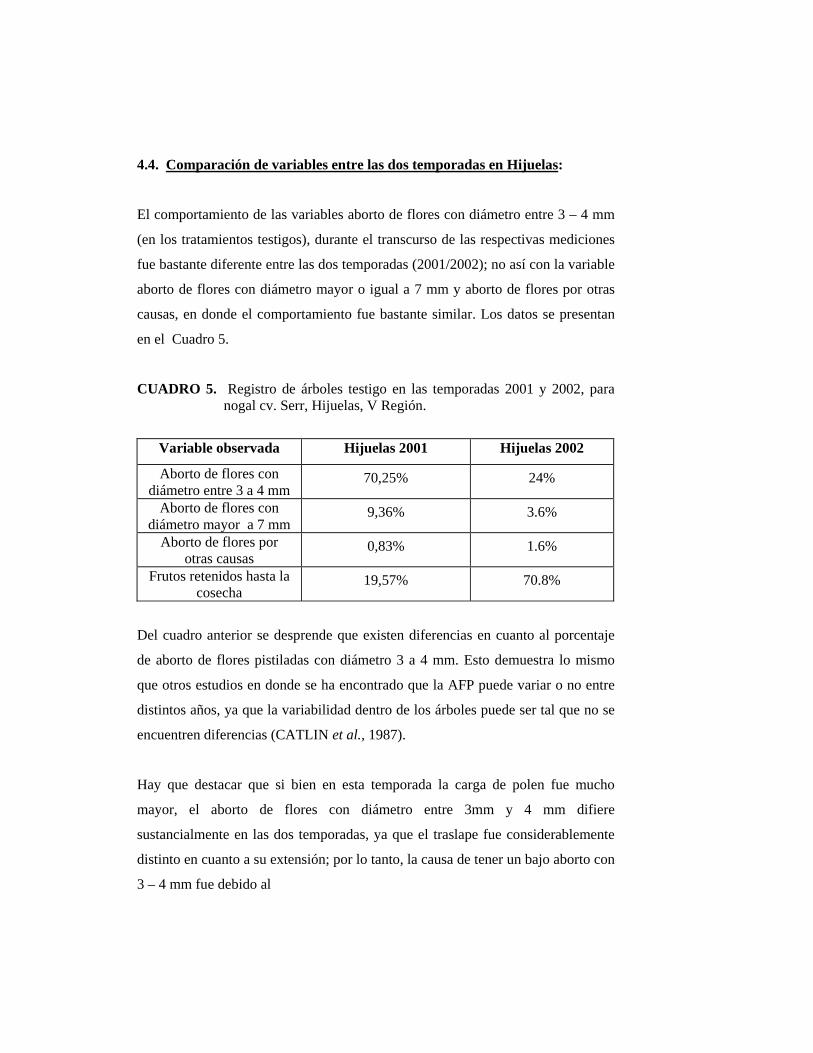
4.4. Comparación de variables entre las dos temporadas en Hijuelas:
El comportamiento de las variables aborto de flores con diámetro entre 3 – 4 mm
(en los tratamientos testigos), durante el transcurso de las respectivas mediciones
fue bastante diferente entre las dos temporadas (2001/2002); no así con la variable
aborto de flores con diámetro mayor o igual a 7 mm y aborto de flores por otras
causas, en donde el comportamiento fue bastante similar. Los datos se presentan
en el Cuadro 5.
CUADRO 5. Registro de árboles testigo en las temporadas 2001 y 2002, para nogal cv. Serr, Hijuelas, V Región.
Variable observada Hijuelas 2001 Hijuelas 2002
Aborto de flores con diámetro entre 3 a 4 mm
70,25% 24%
Aborto de flores con diámetro mayor a 7 mm
9,36% 3.6%
Aborto de flores por otras causas
0,83% 1.6%
Frutos retenidos hasta la cosecha
19,57% 70.8%
Del cuadro anterior se desprende que existen diferencias en cuanto al porcentaje
de aborto de flores pistiladas con diámetro 3 a 4 mm. Esto demuestra lo mismo
que otros estudios en donde se ha encontrado que la AFP puede variar o no entre
distintos años, ya que la variabilidad dentro de los árboles puede ser tal que no se
encuentren diferencias (CATLIN et al., 1987).
Hay que destacar que si bien en esta temporada la carga de polen fue mucho
mayor, el aborto de flores con diámetro entre 3mm y 4 mm difiere
sustancialmente en las dos temporadas, ya que el traslape fue considerablemente
distinto en cuanto a su extensión; por lo tanto, la causa de tener un bajo aborto con
3 – 4 mm fue debido al
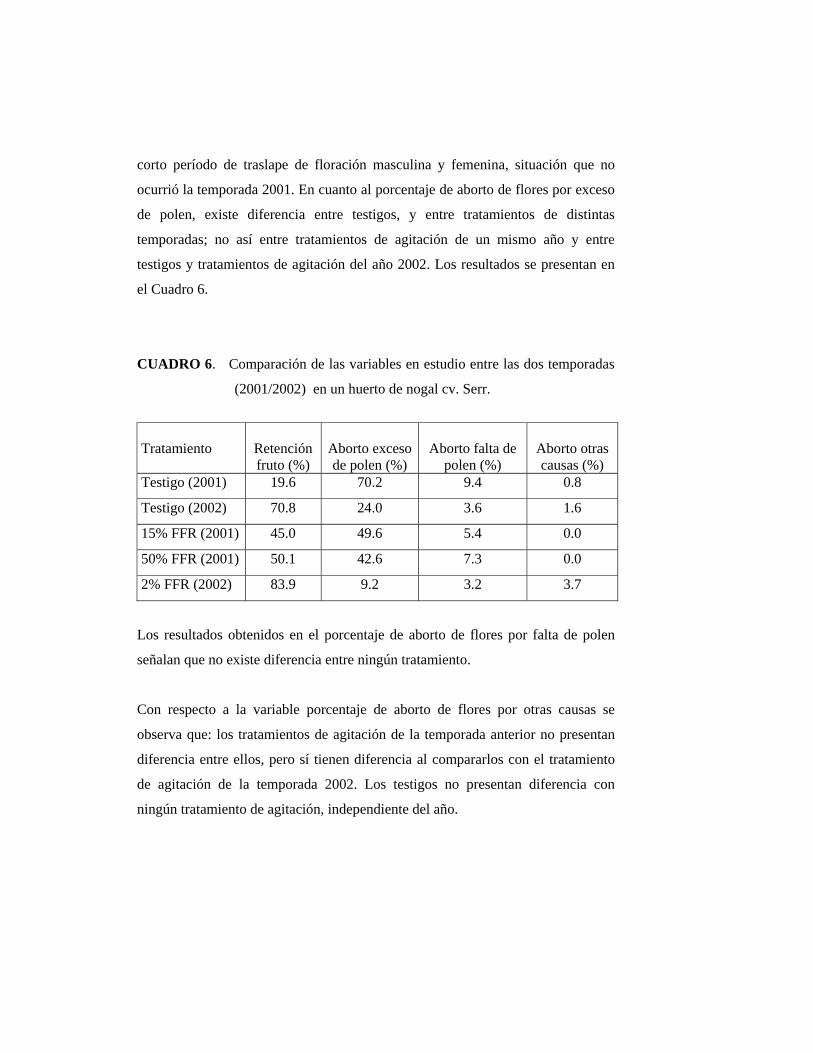
corto período de traslape de floración masculina y femenina, situación que no
ocurrió la temporada 2001. En cuanto al porcentaje de aborto de flores por exceso
de polen, existe diferencia entre testigos, y entre tratamientos de distintas
temporadas; no así entre tratamientos de agitación de un mismo año y entre
testigos y tratamientos de agitación del año 2002. Los resultados se presentan en
el Cuadro 6.
CUADRO 6. Comparación de las variables en estudio entre las dos temporadas
(2001/2002) en un huerto de nogal cv. Serr.
Los resultados obtenidos en el porcentaje de aborto de flores por falta de polen
señalan que no existe diferencia entre ningún tratamiento.
Con respecto a la variable porcentaje de aborto de flores por otras causas se
observa que: los tratamientos de agitación de la temporada anterior no presentan
diferencia entre ellos, pero sí tienen diferencia al compararlos con el tratamiento
de agitación de la temporada 2002. Los testigos no presentan diferencia con
ningún tratamiento de agitación, independiente del año.
Tratamiento
Retención fruto (%)
Aborto exceso de polen (%)
Aborto falta de
polen (%)
Aborto otras causas (%)
Testigo (2001) 19.6 70.2 9.4 0.8
Testigo (2002) 70.8 24.0 3.6 1.6
15% FFR (2001) 45.0 49.6 5.4 0.0
50% FFR (2001) 50.1 42.6 7.3 0.0
2% FFR (2002) 83.9 9.2 3.2 3.7

4.5. Análisis de registros climáticos :
Durante el período de 29 días que abarcó la floración masculina y femenina se
registró una temperatura media promedio de 12.6°C, teniendo una temperatura
mínima media de 5.3°C y una máxima media de 10.3 °C. Sin embargo, no
existieron grandes fluctuaciones (Figura 14). De esto se puede inferir que los días
fueron más bien calurosos que fríos. Según GIL-ALBERT (1998) con
temperaturas entre 6° a 25°C la floración se desarrolla en forma óptima.
La humedad relativa media registrada durante el período completo de floración
fue de 74.5%, teniendo una máxima media de 86% y una mínima media de 56%.
De lo anterior se puede inferir que la floración se desarrolló bajo un ambiente más
bien húmedo, lo que podría explicar la presencia de peste negra en el huerto, lo
que hizo aumentar el porcentaje de aborto de flores pistiladas (PINILLA y
ALVAREZ, 1997).
Por otra parte, WESTWOOD (1993) señala que el clima influye en la dicogamia
durante los períodos cálidos y los amentos responden más rápidamente que la flor
femenina, haciendo mayor la dicogamia en cultivares protoándricos. Además,
señala que la temperatura y la humedad tienen un efecto directo en la extensión de
la floración; días calurosos y secos acortan el período de floración. De acuerdo
con el Ensayo realizado la temporada 2002 en Hijuelas, el corto período de
floración fue influenciado directamente por la temperatura.
La emergencia de los amentos y la diseminación de polen aumenta o se acelera
con temperaturas elevadas y está asociada a una humedad relativa baja.
Probablemente el factor más importante que controla la emergencia de las flores
pistiladas y su receptividad es la temperatura, a través de su efecto directo o
indirecto sobre la elongación de la rama y hoja (CECICH, 2003).
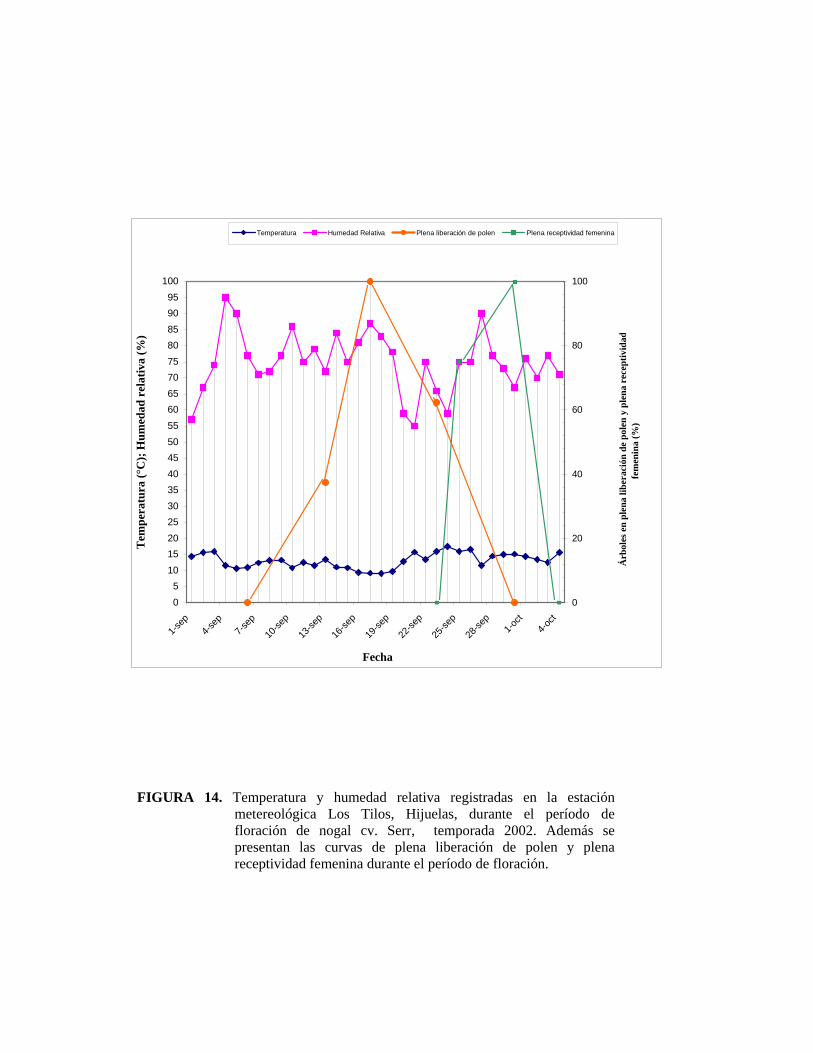
FIGURA 14. Temperatura y humedad relativa registradas en la estación
metereológica Los Tilos, Hijuelas, durante el período de floración de nogal cv. Serr, temporada 2002. Además se presentan las curvas de plena liberación de polen y plena receptividad femenina durante el período de floración.
05
101520253035404550556065707580859095
100
1-sep
4-sep
7-sep
10-se
p
13-se
p
16-se
p
19-se
p
22-se
p
25-se
p
28-se
p1-o
ct4-o
ct
Fecha
Tem
pera
tura
(°C
); H
umed
ad r
elat
iva
(%)
0
20
40
60
80
100
Árb
oles
en
plen
a lib
erac
ión
de p
olen
y p
lena
rec
eptiv
idad
fem
enin
a (%
)
Temperatura Humedad Relativa Plena liberación de polen Plena receptividad femenina

La proporción de los sexos puede variar con las condiciones del lugar. La
variación en temperatura en un gradiente puede influir la base fisiológica para la
asignación de los sexos (CECICH, 2003). Con un aumento de temperatura existirá
un aumento del sexo masculina en el cv. Serr (WESWOOD, 1993).
Al analizar el registro de la precipitación desde el 19 de julio hasta el 22 de
diciembre, se observó que durante el peak de liberación de polen se presentó un
evento de lluvia de 16 mm, lo que no afectó la cuaja de la fruta (Figura 15).
El viento es otro factor climático que puede afectar seriamente a las plantaciones
frutales (FERNÁNDEZ, 1996). Influye directamente en la rotura de ramas, caída
y daño de frutos, dificulta la formación de árboles jóvenes y tiene gran
importancia en la polinización de algunos huertos frutales. La velocidad del viento
promedio entre los meses de agosto a noviembre del 2002 fue de 3.15 mseg-1, lo
que corresponde a una brisa ligera (GRACE, 1977). Con esta velocidad de viento
la dispersión de granos de polen ocurrió en todo el huerto, ya que el polen al ser
liviano le bastan velocidades del orden de 1 mseg-1 para ser transportado
(POLITO, 1998). Se registró una máxima promedio en el mes de noviembre de
6.4 mseg-1 (Figura 16).
FIGURA 15. Precipitación (mm) durante el período de floración (Estación metereológica Los Tilos, Hijuelas). Temporada 2002.
0
10
20
30
40
50
60
70
80
90
100
19/7
27/7 4/8 12
/820
/828
/8 5/9 13/9
21/9
29/9
7/10
15/10
23/10
31/10 8/1
116
/1124
/11 2/12
10/12
Fecha
Prec
ipita
ción
(mm
)
0
10
20
30
40
50
60
70
80
90
100
Árb
oles
(%)
Precipitación
Plena liberaciónde polen
Plenareceptividadfemenina
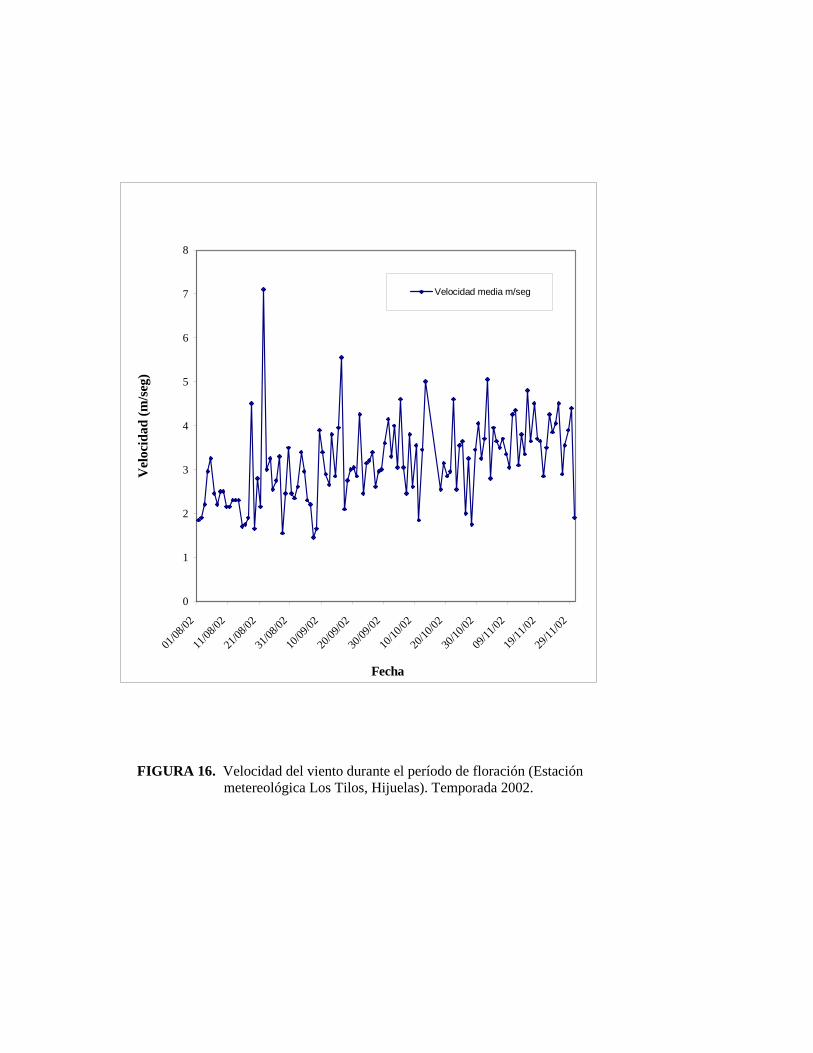
FIGURA 16. Velocidad del viento durante el período de floración (Estación metereológica Los Tilos, Hijuelas). Temporada 2002.
0
1
2
3
4
5
6
7
8
01/08
/02
11/08
/02
21/08
/02
31/08
/02
10/09
/02
20/09
/02
30/09
/02
10/10
/02
20/10
/02
30/10
/02
09/11
/02
19/11
/02
29/11
/02
Fecha
Vel
ocid
ad (m
/seg
)
Velocidad media m/seg

5. CONCLUSIONES
El tratamiento de agitación del árbol de nogal cv. Serr, realizado con un 2% de
flores femeninas receptivas, permite aumentar el rendimiento en un 15%, en
relación al testigo sin agitación.
Se logró determinar que la fenología floral del nogal cultivar Serr en un mismo
huerto difiere de año en año, mostrando diferencias de una semana en la extensión
de la floración, siendo la temporada 2001 más extensa que la temporada 2002, y
de traslape entre floración femenina y masculina, donde la temporada 2001 fue
cubierta durante 18 días, al contrario de esta temporada donde fue cubierta sólo 9
días.
Con respecto a la carga de polen total registrada durante las temporadas 2001 y
2002, se concluye que esta última fue un 38% superior a la registrada el año
anterior. Ahora bien, al momento en que las flores femeninas se encontraban
receptivas, la carga de polen fue menor que la temporada 2001, lo que se tradujo
en un porcentaje de exceso de polen menor, y por ende un bajo porcentaje de
aborto de flores pistiladas.
Se encontró efecto del tratamiento sobre las variables aborto de flores pistiladas
con diámetro entre 3-4 mm y retención de frutos hasta la cosecha. Por lo tanto, el
tratamiento de agitación es efectivo para disminuir el problema del aborto de
flores pistiladas con diámetro entre 3-4 mm y para aumentar la productividad en
nogales cv. Serr para el huerto Los Tilos, V región. No se observó diferencias en
las flores abortadas con diámetro mayor o igual a 7 mm y flores abortadas por
otras causas.

6. RESUMEN
Entre los meses de septiembre de 2002 y febrero de 2003 se llevó a cabo un estudio en un nocedal de la V región (Hijuelas), plantado en 1994, con el objetivo de caracterizar la fenología floral, cuantificar la carga de polen y el aborto de flores pistiladas en el cv. Serr. Se evaluó además el manejo de la polinización mediante agitación de los árboles en un estado fenológico (porcentaje de flores femeninas receptivas menor al 5%) para disminuir el aborto de flores pistiladas y así mejorar la retención de fruta. Se cuantificó el: aborto de flores pistiladas con diámetro de 3-4 mm, aborto de flores con diámetro mayor o igual a 7 mm y aborto de flores por daño de: polilla de la manzana (Cydia pomonella L.) y peste negra del nogal (Xanthomonas campestris pv. Juglandis). Al final de la temporada se cuantificó el número de frutos retenidos en el árbol. Los resultados mostraron en los árboles testigos que la fenología floral masculina se inició dos semanas antes que la floración femenina mostrando una clara diferencia de mayor extensión y mayor traslape con la situación ocurrida durante el año 2001. La carga de polen fue un 38% superior que la registrada en le temporada anterior. El aborto de flores pistiladas con diámetro 3-4 mm por exceso de polen fue de un 24.01% la temporada 2002 y de un 70,25% la temporada 2001, el aborto de flores pistiladas con diámetro mayor o igual a 7 mm por falta de polen fue de 5,10% el año 2002 y de un 9,36% el año 2001. Se registró la caída de flores por daños de enfermedad y plaga; y la fruta retenida hasta la cosecha, siendo de 1,63% y un 70,80%, respectivamente, para el año 2002 y de 0,83% y un 19,5%, respectivamente para el año 2001. El hecho de realizar el tratamiento de agitación de amentos con un 2% de flores femeninas receptivas aumenta la producción del huerto Los Tilos en un 15% en relación a las árboles testigo sin agitación, lo que se puede explicar por una disminución en el aborto de flores pistiladas en un 61,7% en los árboles con agitación.
Comentario: Esto da a entender que el manejo de la polinización se realizó en los dos huertos.

7. LITERATURA CITADA
ARAYA, X. 2000. Estudio fenológico de los principales cultivares de Nogal Europeo en la Región Metropolitana. Tesis Ing. Agr. Santiago, Pontificia Universidad Católica de Chile, Facultad de Agronomía e Ingeniería Forestal. 45 p.
ALETA, N. y NINOT, A. 1993. Variedades de nogal. Fruticultura Profesional 54:
93-100. ASTORGA, B.E., y ASTORGA, M.E. 1987. Peste negra del nogal o bacteriosis
y su control. El Campesino 118 (9): 58-61 BINGGELI, P. 2003. Ratio of protandrous to protogynous individuals and the
internase of these characteristics, (on line). http://www.members.lycos.co.uk/woodyplantecology/sycamore/ratio.htm.
CAMPOS, A. y VALDERRAMA, E. 2001. Mercado. In: Lemus, G., ed. El
nogal en Chile. Santiago, INIA. pp. 207 – 224 CARREÑO, I. y PINTO, A. 1988. Control químico de peste negra del nogal.
Agricultura Técnica 48 (3): 258-261. CATLIN, P. 1998. Root physiology and rootstocks. In: Walnut Production
Manual. California, University of California. pp: 35-46. (Publication 3373).
________. and OLSSON, E. 1990. Pistillate Flower Abscission of Walnut - Serr,
Sunland, Howard, and Chandler. HortScience 25 (11): 1391 - 1392. ________. and POLITO, V. 1989. Cell and tissue damage associated with
pistallate flower abscission of Persian Walnut. HortScience 24 (6): 1003 - 1005.
________., RAMOS, D., SIBBETT, G., OLSSON, W. and OLSSON, E. 1987.
Pistillate flower abscission of persian walnut. HortScience (22): 201-205.
CECICH, R. 2003. Floral biology and pollination of Eastern black walnut, (on
line). http://www.ncrs.fs.fed.us/epubs/walnut/p79_89.pdf CENTRO DE INFORMACIÓN DE RECURSOS NATURALES –
CORPORACIÓN DE FOMENTO PARA LA PRODUCCION. 1997. Estudio Agrológico, V Región. Santiago, CIREN. 180 p.

CHARLOT, G. et GERMAIN, E. 1988. Le noyer nouvelles techniques. France, Edition CTIFL. 208 p.
DENG, X., WEINBAUM, S., DEJONG, T. and MURAOKA, T. 1991. Pistillate
Flower Abortion in Serr Walnut associated with reduced carbohydrate and nitrogen concentrations in wood and xylem sap. Journal of the American Society for Horticultural Science. 116 (2): 291-296.
ESTAY, P. 1997. Principales plagas del nogal en Chile. In: Seminario Avances
Tecnológicos en el Cultivo del Nogal. Santiago, INIA. pp. 96-106 ESTERIO, M. 1984. Bacteriosis del nogal. Aconex 7 : 21-26 FERNANDEZ, R. 1996. Planificación y diseño de plantaciones frutales.
Madrid. Ediciones Mundiprensa. 220 p. FORDE, H. and GRIGGS, W. 1981. Pollination and blooming habits of walnuts.
California, Division of Agriculture and Natural Resources cooperative. University of California. 4 p. (Lflt. 2753).
__________. and __________. 1975. Pollination and blooming habits of
walnuts. California, Division of Agriculture and Natural Resources cooperative. University of California. (Lflt. 2753).
__________. and __________. 1972. Pollination and blooming habits of walnuts.
Univ. Of California. (AXT 24). GERMAIN, E., JALINAT, J. et MARCHOU, M. 1975. Divers aspects de la
biologie florale du noyer. (Juglans regia L.) Extract from Bulletin Technique d’ information. INVUFLEC. 282: 13-14
GIL, G. 2000. La producción de fruta. Fruta de climas templados y subtropical y
uva de vino. Santiago, Editorial Universidad Católica de Chile. 583 p.
GIL-ALBERT, F. 1996. Tratado de arboricultura frutal. Morfología y fisiología
del árbol frutal. Madrid, Editorial Mundiprensa. 102 p. (Vol 1). HENDRICKS, L., COATES, W., ELKINS, R., McGRANAHAN, G., PHILLIPS,
H., RAMOS, D., REIL, W. and SNYDER, R. 1998. Selection of varieties. In: Walnut Production Manual. California, University of California. pp: 84 - 89. (Publication 3373).

HERRERA, R. 2002. Caracterización del aborto de flores pistiladas en nogal cv. Serr y manejo de la polinización para mejorar la retención de fruta, en dos localidades de la V región. Taller de Licenciatura. Agr. Quillota, Universidad Católica de Valparaíso, Facultad de Agronomía. 73 p.
HUDSON, T. WILLIAM, J. and ANTOM, M. 1981. Growth, Development, and
Utilization of Cultivated Plants. New Jersey, Prentice-Hall. 606-607 pp.
KRUEGER, W. H. 2000. Pollination of english walnuts: Practices and problems.
HortTechnology 10 (1): 127 – 130 LATORRE, B., y ESTERIO, M. 1981. Peste negra del nogal. El Campesino, 112
(7): 16-18. LEMUS, G., VALENZUELA, J. y LOBATO, A. 2001. Origen y botánica. In:
Lemus, G. ed. El nogal en Chile. Santiago, INIA. pp: 9-23. LIN, J., SHABNAY B. and RAMOS D. 1977. Pistillate flower development and
fruit gwoth in somo English walnut cultivars. Journal of the American Society for Horticultural Science. 102: 702-705.
LUNA, F. 1990. El nogal. Producción de fruta y madera. Madrid, Ediciones
Mundi-Prensa. 155 p. LUZA, J. and POLITO V. 1988. Microsporogenesis and another differentiation
in Juglans regia L.: a developmental basis for heterodichogamy. Bot anical Gazette. 149: 30-36.
McGRANAHAN, G., VOYIATZIS, D., CATLIN, P. and POLITO, V. 1994. High
pollen load can cause pistillate flower absicission in walnut. Journal of the American Society for Horticultural Science. 199 (3): 505 - 509.
MINISTERIO DE AGRICULTURA, OFICINA DE ESTUDIOS Y POLITICAS
AGRARIAS. 2003. Condiciones de accesos de los productos silvoagropecuarios chilenos en los mercados externos, (online). http://www.odepa.gob.cl/internacional/ConAcceso/CAcceso99.pdf
_________. 2001. Compendio estadístico silvoagropecuario 1990 – 2000.
Santiago, Chile, ODEPA. 172 p. MUNCHARAZ, M. 2001. El nogal. Técnicas de cultivo para la producción
frutal. Madrid, Mundi - Prensa. 300 p.

PASTORE, P. 2003. El cultivo de las nueces, (on line). http://www.ambiente-ecológico.com/ediciones/2002/085_09.2002/085_Flora_Infoagro.php3
PINNEY, K., LABAVITCH, J. and POLITO, V. 1998. Fruit growth and
development. In: Walnut Production Manual. California, University of California. pp: 139-143. (Publication 3373).
PINILLA, B. y ALVAREZ, M. 1997. Principales enfermedades del nogal en
Chile. In: Seminario “Avances tecnológicos en el cultivo del nogal”. CRI La Platina. Santiago, 12-14 noviembre, 1997. pp: 90-95.
POLITO, V. 1998. Floral Biology: Flower structure, development and
pollinitation. In: Walnut Production Manual. California, University of California. pp: 127 -132. (Publication 3373).
________. and N. Y. Li. 1985. Pistillate flower differentation in English walnut (Juglans regia L.): developmental basis for heterodichogamy. Scientia Horticulturae 26: 333-338.
________. and PINNEY, K. 1997. The relationship between phenology of
pistillate flower organogenesis and mode of heterodichogamy in Juglans regia L. (Juglandaceae). Sexual Plant Reproduction 10 (1): 36-39
________., SIBBETT, G., GRANT, J., KELLEY, K. and CATLIN, P. 1998.
Pistillate flower abortion and pollination management. In: Walnut Production Manual. California, University of California. pp: 133 - 138. (Publication 3373).
POR, A. and POR, J. 1990. The effect of the excess of pollen on the fruitset of
walnuts in Balaton-boglár. Acta Horticulturae 284: 253 – 257. PRADO, E. 1998. Plagas del nogal. In: 2º Curso de producción de nogal INIA –
La Platina. San Esteban, 19 y 20 de agosto de 1998. pp: 101-111. RENOVALES, G. 2001. Plantas de interés farmacéutico, 72 especies presentes
en la península ibérica, (on line). http://www.vc.ehu.es/plfarm/18.jure.htm
ROVIRA, M., NINOT, A. and ALETÀ, N. 2001. Pistillate flower abortion in
walnut (J. regia L.). Acta Horticulturae. 544: 287 - 293 ________. and ALETÀ, N. 1997. Pistillate flower abscission on four walnut
cultivars. Acta Horticulturae. 442: 231 – 234

RYUGO, K. 1986. Promotion and inhibition of flower and fruit set by plant manipulation and hormones. Acta Horticulturae. 179: 301 - 307
________., BARTOLINI, G., CARLSON, R. M. and RAMOS, D. E. 1985.
Relationship between catkin development and cropping in the Persian walnut “Serr”. Hortscience 20 (6): 1094 – 1096
SANTIBAÑEZ, F. y URIBE, J. 1990. Atlas Agroclimático de Chile. Santiago,
Universidad de Chile. 65 p. SIBBETT, G., COATES, W. and EDSTROM, J. 1998. Orchard planning, desing
and planting. In: Walnut Production Manual. California, University of California. pp: 90-98. (Publication 3373).
VALENZUELA, J. , LOBATO, A. y LEMUS, G. 2001. Cultivares. In: Lemus, G.
ed. El nogal en Chile. Santiago, INIA. pp: 41-51. ________., ESPINOZA, J. y PARRA, A. 1992. Variedades comerciales de nogal.
Santiago, Instituto de Investigaciones Agropecuarias. Serie La Platina Nº 34.7
WESTWOOD, M. 1993. Temperate Zone Pomology: Physiology and Culture.
San Francisco, Freeman. 523 p.

ANEXOS

ANEXO 1a: Tiempo (semanas) de los estados más importantes en el desarrollo del fruto del nogal.
Semanas después de
floración
Etapa de desarrollo
1 Fertilización de la célula huevo
2 Embrión consistente en 2 a 8 células
3 Embrión consistente en 8 a 32 células
5 Embrión globular y consta de varios cientos de células
6 Los cotiledones comienzan su crecimiento. Comienza
el endurecimiento de la cáscara en el ápice.
8 El crecimiento comienza a disminuir en comparación
a un estado más temprano y rápido.
9 La carne dela nuez comienza un rápido crecimiento
mientras los cotiledones comienzan a llenar los
lóculos.
10 Tamaño final de la nuez
12-15 La cáscara completa su esclerificación
15 Se reanuda el crecimiento de la carne de la nuez con
rapidez.
18 Peso total máximo
19-22 Fecha aproximada de cosecha.
Fuente: Polito, 1998
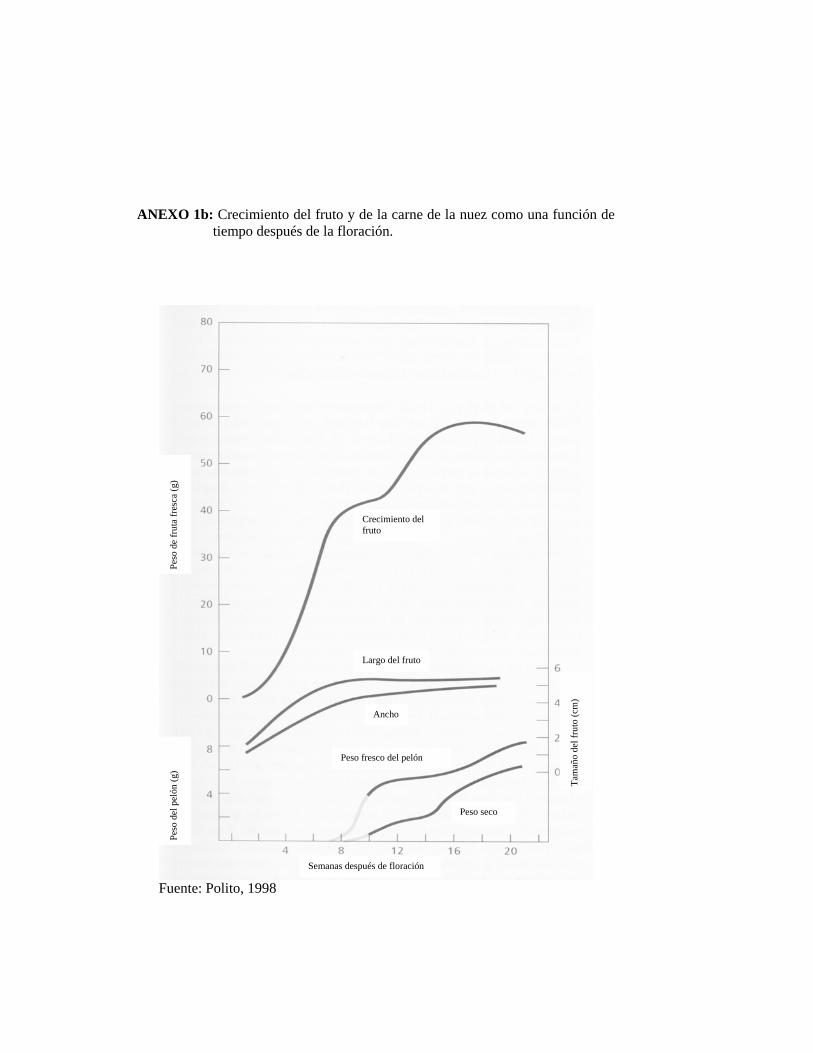
ANEXO 1b: Crecimiento del fruto y de la carne de la nuez como una función de
tiempo después de la floración.
Fuente: Polito, 1998
Peso
de
frut
a fr
esca
(g)
Crecimiento del fruto
Peso
del
pel
ón (g
)
Largo del fruto
Tam
año
del f
ruto
(cm
)
Ancho
Peso fresco del pelón
Peso seco
Semanas después de floración

ANEXO 2: Labores culturales desarrolladas en Los Tilos, Hijuelas, durante la temporada 2002.
1- Poda: se efectúo en todos los árboles buscando principalmente la entrada de
luz tomando en cuenta la posición del sol. La iluminación es lo más buscado,
sobre todo en las variedades muy vigorosas. Para esto se sacaron ramas de
mediano grosor y cualquier tipo de ramificación vegetativa que no permita una
buena iluminación de la copa, tal como chupones y ramas mal colocadas o
distribuidas. Además se sacaron ramillas débiles y secas. De esta manera se buscó
principalmente hacer un aclareo de ramas manteniendo en nivel y la calidad
productiva. También se hizo un levantamiento de faldas a todos los árboles.
2.- Control de Phytophtora cactorum: se hizo un manejo a todas las plantas del
huerto, aunque las únicas que se encontraban dañadas por el hongo eran aquellas
injertadas sobre el patrón Juglans nigra, debido a que este patrón es el más
susceptible a esta enfermedad. El manejo fue hacer un tazón amplio, exponiendo
la inserción de las raíces próximas a la superficie del suelo. En la base de los
troncos se colocó cloro puro y con brocha se aplicó una solución de Ridomil más
agua en dosis de 250 grL-1 de agua.
3.- Control de malezas: se hizo en dos épocas, la primera fue en invierno
aplicando Gesatop (Simazina 90%, nombre comercial Gesatop90) en dosis de 24
gL-1 de agua. Durante la primavera se aplicó glifosato en dosis de 12 mLL-1 de
agua y por otro lado se aplicó amizol usando 24 mLL-1 de agua.
Las aplicaciones se hicieron durante la mañana, tratando de evitar las horas de
mayor calor y el viento. Se aplicó sólo a la entre hilera, dejando en la sobrehilera
las malezas sin control.
4.- Control plagas y enfermedades: A mediados de Diciembre se hizo una
aplicación contra la polilla del nogal. Vía aérea se aplicó Gusathion (Azinphos-
metyl, nombre comercial Gusation M 35%) en dosis de 62500 mLL-1 de agua /
ha.
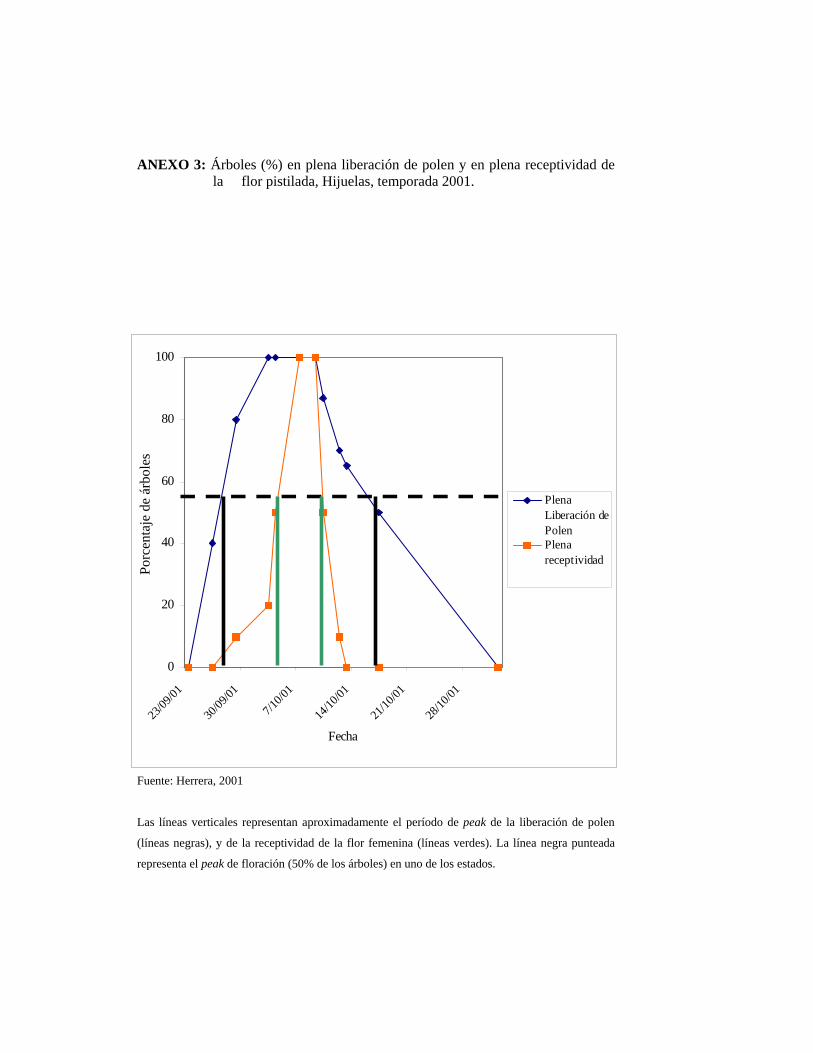
ANEXO 3: Árboles (%) en plena liberación de polen y en plena receptividad de la flor pistilada, Hijuelas, temporada 2001.
Fuente: Herrera, 2001
Las líneas verticales representan aproximadamente el período de peak de la liberación de polen
(líneas negras), y de la receptividad de la flor femenina (líneas verdes). La línea negra punteada
representa el peak de floración (50% de los árboles) en uno de los estados.
0
20
40
60
80
100
23/09
/01
30/09
/01
7/10/0
1
14/10
/01
21/10
/01
28/10
/01
Fecha
Porc
enta
je d
e ár
bole
s
PlenaLiberación dePolenPlenareceptividad
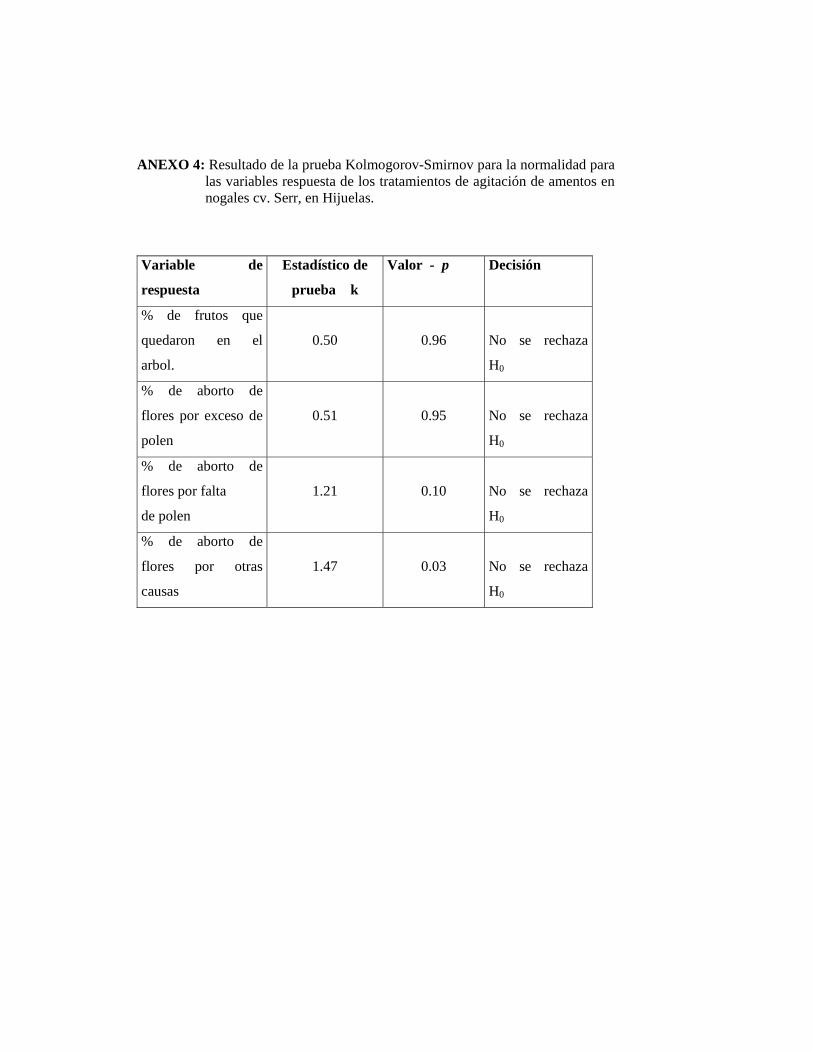
ANEXO 4: Resultado de la prueba Kolmogorov-Smirnov para la normalidad para las variables respuesta de los tratamientos de agitación de amentos en nogales cv. Serr, en Hijuelas.
Variable de
respuesta
Estadístico de
prueba k
Valor - p Decisión
% de frutos que
quedaron en el
arbol.
0.50
0.96
No se rechaza
H0
% de aborto de
flores por exceso de
polen
0.51
0.95
No se rechaza
H0
% de aborto de
flores por falta
de polen
1.21
0.10
No se rechaza
H0
% de aborto de
flores por otras
causas
1.47
0.03
No se rechaza
H0
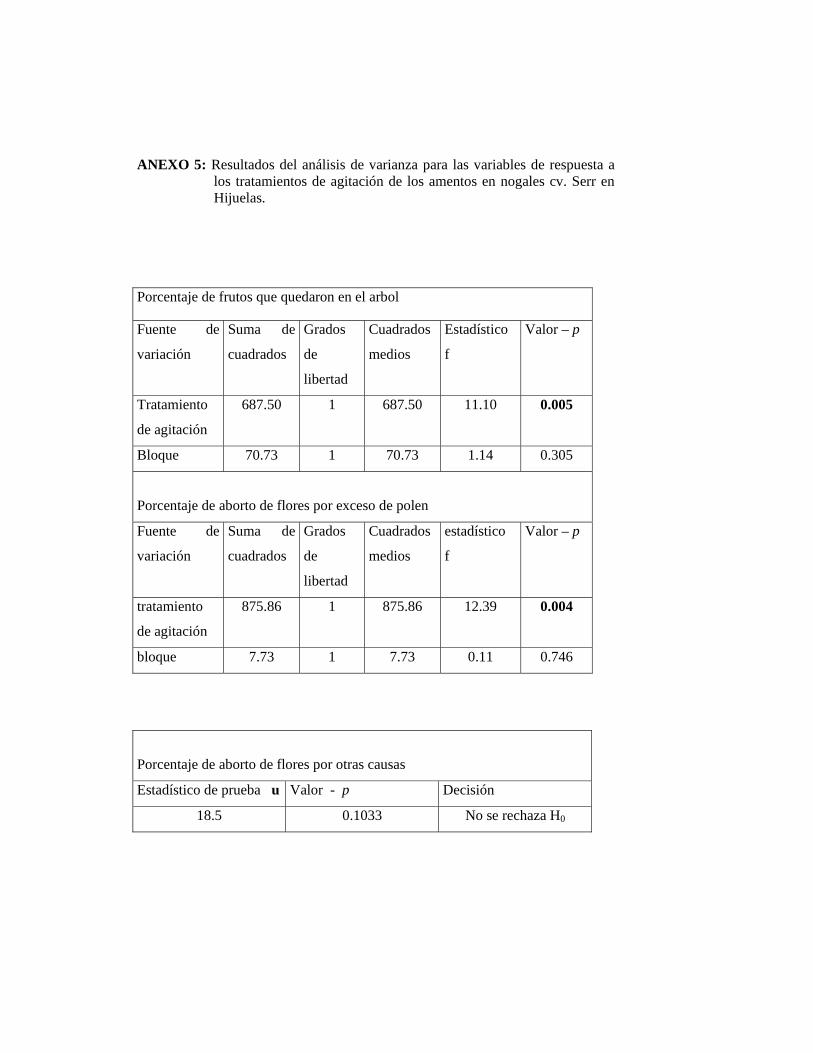
ANEXO 5: Resultados del análisis de varianza para las variables de respuesta a los tratamientos de agitación de los amentos en nogales cv. Serr en Hijuelas.
Porcentaje de frutos que quedaron en el arbol
Fuente de
variación
Suma de
cuadrados
Grados
de
libertad
Cuadrados
medios
Estadístico
f
Valor – p
Tratamiento
de agitación
687.50 1 687.50 11.10 0.005
Bloque 70.73 1 70.73 1.14 0.305
Porcentaje de aborto de flores por exceso de polen
Fuente de
variación
Suma de
cuadrados
Grados
de
libertad
Cuadrados
medios
estadístico
f
Valor – p
tratamiento
de agitación
875.86 1 875.86 12.39 0.004
bloque 7.73 1 7.73 0.11 0.746
Porcentaje de aborto de flores por otras causas
Estadístico de prueba u Valor - p Decisión
18.5 0.1033 No se rechaza H0

Porcentaje de aborto de flores por falta de polen
Fuente de
variación
Suma de
cuadrados
Grados
de
libertad
Cuadrados
medios
Estadístico
f
Valor – p
Tratamiento
de agitación
0.59 1 0.59 0.04 0.839
Bloque 54.13 1 54.13 3.97 0.068
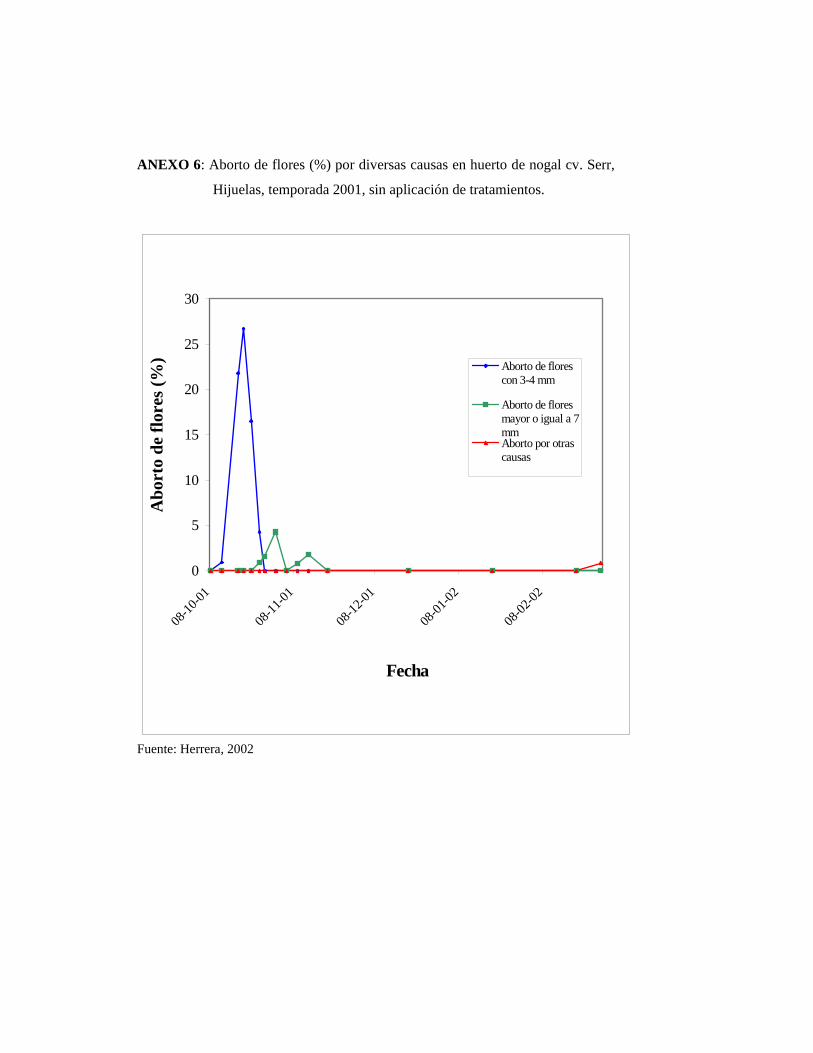
ANEXO 6: Aborto de flores (%) por diversas causas en huerto de nogal cv. Serr,
Hijuelas, temporada 2001, sin aplicación de tratamientos.
Fuente: Herrera, 2002
0
5
10
15
20
25
30
08-10
-01
08-11
-01
08-12
-01
08-01
-02
08-02
-02
Fecha
Abo
rto
de fl
ores
(%) Aborto de flores
con 3-4 mm
Aborto de floresmayor o igual a 7mmAborto por otrascausas


















