Dinamica celular
40
G BARQUISIMETO, FEBRERO 2011 UNIVERSIDAD CENTROCCIDENTAL LISANDRO ALVARADO SISTEMA DE EDUCACION A DISTANCIA DECANATO DE CIENCIAS DE LA SALUD CURSO PREUNIVERSITARIO DINAMICA CELULAR CURSO PREUNIVERSITARIO DECANATO DE CIENCIAS DE LA SALUD.
Transcript of Dinamica celular

G
BARQUISIMETO, FEBRERO 2011
UNIVERSIDAD CENTROCCIDENTALLISANDRO ALVARADO
SISTEMA DE EDUCACION A DISTANCIADECANATO DE CIENCIAS DE LA SALUD
CURSO PREUNIVERSITARIO
DINAMICA CELULAR
CURSO PREUNIVERSITARIO
DECANATO DE CIENCIAS DE LA SALUD.

Dra. Aura ChavezDra. María Elena olivares
UNIDAD IIIDINAMICA CELULAR
CURSO PREUNIVERSITARIO
DECANATO DE CIENCIAS DE LA SALUD.
UNIVERSIDAD CENTROCCIDENTALLISANDRO ALVARADO
SISTEMA DE EDUCACION A DISTANCIADECANATO DE CIENCIAS DE LA SALUD
CURSO PREUNIVERSITARIO

3
Unidad III Dinámica Celular
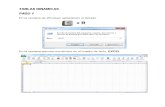
PáginaIntroducción……………………………………………………………………….. 4Objetivo terminal…………………………………………………………………... 4Objetivos específicos……...….……………………………………………………. 4Transporte celular………………….…………………………………………….. 6 Modelo del Mosáico Fluído……..………………………….………………… 6Composición de las membranas…………………………………………………… 7 Proteínas Integrales………………………………………………………….. 7 Proteínas Periféricas…………………………………………………………... 7 Carbohidratos………………………………………………………………… 7 Lípidos……………………………………………………………………….. 7 Movimientos de los lípidos y proteínas en la membrana celular………….. 7 Movimiento Flip-Flop………………………………………………….. 8Fluidez de la membrana…………………………………………………………. 8Permeabilidad de la membrana…………………………………………………. 8Transporte Celular………………………………………………………………. 8
Transporte pasivo…………………………………………………………… 9Transporte activo……………………………………………………………. 9
Endocitosis…………………………………………………………………… 13Actividad Control 1………………………………………………………………. 15Glucolisis…………………………………………………………………………. 16
Glucólisis aeróbica………………………………………………………….. 16Glucólisis anaeróbica……………………………………………………….. 16Fermentación Homoláctica…………………………………………………. 19Fermentación Alcohólica…………………………………………………… 19
Ciclo del Ácido Cítrico…………………………………………………………… 20Cadena Respiratoria o Transportadora de electrones…………………………… 22Mitocondria……………………………………………………………………….. 25Actividad Control 2………………………………………………………………. 27Bibliografía……………………………………………………………………….. 28
TABLA DE CONTENIDOS
Curso Preuniversitario. Biología Celular. Dinámica celular, 2011. Chávez, A y, Olivares, M.

4
Unidad III Dinámica Celular
INTRODUCCIONLas células pueden llevar a cabo múltiples funciones que definen la vida. El
mantenimiento de la vida celular es la expresión de la acción coordinada de los
componentes subcelulares. Muchos de los procesos básicos como el transporte
celular, la síntesis de proteínas, la conversión de energía química son notablemente
similares en los organismos vivos y deben estar debidamente regulados; la alteración
de los mecanismos de regulación celular, se traduce en un desequilibrio celular que
puede conducir hasta la muerte celular. A continuación se introduce al lector en los
procesos celulares básicos de la célula, transporte celular, y transformación de
energía, que permitirán al estudiante aplicar y ampliar los conocimientos abordados
en las unidades precedentes.
Objetivo General
Analizar los mecanismos básicos que rigen el transporte de compuestos a través de la
membrana celular.
Analizar el metabolismo celular.
Objetivos Específicos
1. Identificar los componentes químicos de la membrana celular
2. Clasificar los tipos de transporte celular de acuerdo al gradiente de
concentración.
3. Analizar los mecanismos de transporte utilizados por los compuestos para
atravesar la membrana de acuerdo a la naturaleza química del compuesto.
4. Analizar los mecanismos implicados en el mantenimiento del potencial de
membrana.
5. Comparar los procesos de endocitosis y fagocitosis.
6. Analizar la interrelación entre Glucólisis, Ciclo de los Ácidos Tricarboxílicos
y Cadena Transportadora de electrones
Curso Preuniversitario. Biología Celular. Dinámica celular, 2011. Chávez, A y, Olivares, M.

5
Unidad III Dinámica Celular
Curso Preuniversitario. Biología Celular. Dinámica celular, 2011. Chávez, A y, Olivares, M.

PáginaIntroducción……………………………………………………………………….. 4
Objetivo terminal……………………………………..…………………………... 4Objetivos específicos……...………………………………………………………. 4
Transporte celular………………….…………………………………………….. 6 Modelo del Mosáico Fluído……..………………………….………………… 6Composición de las membranas…………………………………………………… 7 Proteínas Integrales………………………………………………………….. 7 Proteínas Periféricas…………………………………………………………... 7 Carbohidratos………………………………………………………………… 7 Lípidos……………………………………………………………………….. 7 Movimientos de los lípidos y proteínas en la membrana celular………….. 7 Movimiento Flip-Flop………………………………………………….. 8Fluidez de la membrana…………………………………………………………. 8Permeabilidad de la membrana…………………………………………………. 8Transporte Celular………………………………………………………………. 8
Transporte pasivo…………………………………………………………… 9Difusión simple……………………………………………………………. 9Difusión facilitada…………………………………………………………… 9
Transporte activo……………………………………………………………. 9Transporte activo primario. Bomba Na+-K+……………………………… 10Potencial de membrana, depolarización y potencial de acción………….. 11Transporte activo secundario……………………………………………… 12
Endocitosis…………………………………………………………………… 13Fagocitosis…………………………………………………………………. 13Endocitosis mediada por receptores……………………………………… 13
Actividad Control 1………………………………………………………………. 15Glucolisis…………………………………………………………………………. 16
Glucólisis aeróbica………………………………………………………….. 16Glucólisis anaeróbica……………………………………………………….. 16Fermentación Homoláctica…………………………………………………. 19Fermentación Alcohólica…………………………………………………… 19
Ciclo del Ácido Cítrico…………………………………………………………… 20Cadena Respiratoria o Transportadora de electrones…………………………… 22
Fosforilación unida a sustrato………………………………………………….. 22Fosforilación oxidativa………………………………………………………… 23
Mitocondria……………………………………………………………………….. 25Actividad Control 2………………………………………………………………. 27Bibliografía……………………………………………………………………….. 28
CONTENIDO

Transporte CelularLas células están separadas del
ambiente externo por una estructura
denominada membrana celular o
plasmática con un espesor de 5 a 10
nm. A finales del año 1950, con la
ayuda del microscopio electrónico, se
pudo observar la membrana plasmática
como una estructura compuesta por
dos capas de color oscuro orientadas
paralelamente, separadas por una capa
clara (Fig. 1). Este patrón trilaminar, se
repitió en todas las membranas
celulares examinadas, tales como las
plasmáticas, envolturas nucleares, o
citoplasmáticas.
Figura 1. Patrón trilaminar de la membrana plasmática.http://3.bp.blogspot.com/_vwpOdFPkZ6o/TFIK1KvmDvI/AAAAAAAAAMY/l7aAeqjS_gI/s1600/4%C2%BA+Membrana+foto+y+esquema.JPG
Módelo del Mosaico Fluido. Los
hallazgos sobre la membrana,
generaron una serie de debates acerca
de la composición molecular de las
diferentes capas de una membrana, que
llevó a la propuesta de diversos
modelos. Los científicos
estadounidenses Garth Nicolson y
Seymour J. Singer en 1972
propusieron el modelo del mosaico
fluido. Este modelo, mantiene la
disposición de los lípidos en bicapa,
considera que las proteínas se
presentan como un “mosaico” de
partículas discontinuas que penetran y
atraviesan la bicapa lipídica (Fig 2)
Lo más resaltante, del modelo
del mosaico fluido, es considerar las
membranas celulares como estructuras
dinámicas donde sus componentes son
móviles, capaces de interaccionar de
acuerdo a los requerimiento
funcionales de la célula
Figura 2. Modelo de Mosaico Fluido para las membranas biológicashttp://web.usal.es/~rororo/pract2/imagenes/ch12-3.jpg

Composición de las Membranas.
Las membranas biológicas son bicapas
lipídicas en las que se insertan las
proteínas. Los lípidos y proteínas
pueden ser modificados por
carbohidratos constituyendo los
glicolípidos y glicoproteínas.
. Las proteínas pueden ser:
integrales y periféricas. Las
proteínas integrales atraviesan la
membrana y las proteínas periféricas
están unidas a proteínas integrales.
Los carbohidratos se orientan hacia el
exterior celular.
Los lípidos representan
aproximadamente 50% de la
composición de las membranas,
porcentaje que varía de acuerdo al tipo
y funciones de la membrana. Por
ejemplo la membrana interna de la
mitocondria contiene
aproximadamente 70% de proteínas,
organizadas en complejos implicados
en el transporte de electrones y
fosforilación oxidativa.
Las membranas plasmáticas de
mamíferos contienen
aproximadamente de 50% a 60% de
lípidos. Tal como se analizó en la
Unidad Composición Química de la
Célula, entre estos lípidos, se
encuentra: fosfatidil colina, fosfatidil
serina, fosfatidiletanolamina y
esfingomielina. Entre estos lípidos
alternan moléculas de colesterol. A
esta bicapa de lípidos de asocian
proteínas que participan como
receptores de señales externas y
transportadores de compuestos.
Movimientos de los Lípidos y
Proteínas en la Membrana Celular.
En las bicapas lipídicas los lípidos y
las proteínas pueden rotar y moverse
en sentido lateral. Además, los lípidos
pueden moverse de una mitad de la
bicapa a la otra por un movimiento
conocido como flip-flop (Fig. 3).
Figura 3. Movimiento de los lípidos de membranahttp://4.bp.blogspot.com/_PMVRCa6NLO4/TJFBEu_WoVI/AAAAAAAAATU/wPALLSxSooo/s320/fluidez+de+membrana.png
Fluidez de la Membrana. La fluidez
de la membrana está determinada por
la temperatura y por la composición

lipídica (largo de la cadena de los
ácidos grasos y la presencia de dobles
enlaces) tal como se analizó en la
unidad composición química. El
colesterol, también regula la fluidez de
la membrana. Por un lado, la
interacción de los anillos
hidrocarbonados rígidos con las
cadenas hidrocarbonadas de los ácidos
grasos, disminuyen la movilidad de las
porciones externas de los ácidos
grasos, haciendo que esta parte de la
membrana sea más rígida. a
temperatura altas. Por otro lado, al
interferir en las interacciones entre las
cadenas hidrocarbonadas de los ácidos
grasos, mantiene la fluidez de la
membrana a temperaturas más bajas.
Permeabilidad de la
Membrana Celular. La
permeabilidad selectiva de las
membranas biológicas, le permite a las
células controlar y mantener su
composición química.
La permeabilidad de las
membranas, entendida como una
medida de la facilidad con que un
compuesto atraviesa la membrana, una
vez que se requiera, depende de la
composición química de la membrana
y de la naturaleza química del
compuesto.
Transporte Celular. Se
distinguen dos tipos de transporte,
atendiendo al gradiente de
concentración, a ambos lados de la
membrana, transporte pasivo y
transporte activo.
Figura 4. Tipos de transporte de membrana.http://elprofedebiolo.blogspot.com/2010/02/transporte-de-membrana.html
Transporte Pasivo. Se produce
cuando un compuesto difunde a través
de la membrana a favor del gradiente
de concentración; es decir, desde
donde el compuesto está en mayor
concentración hacia donde se
encuentra en menor concentración
(Fig. 4). Dentro del transporte pasivo,
se distinguen la difusión simple y la
difusión facilitada.
Difusión simple: Este tipo de
transporte es experimentado sólo por
moléculas pequeñas no cargadas tales
como glicerol, O2, CO2, H20 y etanol,

que pueden difundir libremente a
través de la membrana.
Difusión facilitada. Se produce
cuando las moléculas polares pequeñas
como los iones, atraviesan la
membrana, a través de proteínas
transmembrana, llamadas proteínas
canal, permitiendo a la célula
controlar el movimiento de iones a
través de la membrana. sin
interaccionar con las cadenas
hidrófobas de los lípidos de la
membrana. Los poros formados por
estos canales proteicos se abren o
cierran selectivamente en respuesta a
señales extracelulares.
Las moléculas pequeñas como
glucosa y aminoácidos pueden
atravesar la membrana por difusión
facilitada, a través de proteínas
transmembrana, llamadas proteínas
transportadoras.
Transporte Activo. Se establece
cuando un compuesto atraviesa la
membrana en contra del gradiente de
concentración; es decir, desde donde
está menos concentrado hacia donde
está más concentrado. Éste tipo de
transporte se divide a su vez en
primario y secundario. El transporte
activo primario está acoplado
directamente a la hidrólisis de ATP y
el transporte activo secundario, está
indirectamente acoplado a la hidrólisis
de ATP.
Al igual que en la difusión
facilitada, el transporte activo de un
compuesto requiere de proteínas
integrales de membrana, conocidas
como proteínas transportadoras. En
este caso, la proteína se une
selectivamente a un compuesto
particular y permite su desplazamiento
a través de la membrana y posterior
liberación, gracias a cambios
conformacionales de la proteína
acoplados a la utilización de energía
almacenada en el ATP.
Por lo general la composición
iónica específica del citosol difiere
apreciablemente del líquido
extracelular circulante. En las células
animales la concentración citosólica de
K+ es 20 a 40 veces mayor en las
células que en la sangre, mientras que
la concentración de Na+ es de 8 a 12
veces menor en las células que en la
sangre. La concentración de Ca+ libre
en el citosol suele ser menor de 0.2

mM, mil o más veces menor que la de
la sangre.
Transporte activo primario.
Bomba Na+-K+. Las bombas de
iones son en gran medida las
encargadas de establecer y mantener
los gradientes iónicos a través de la
membrana plasmática, mediante un
transporte activo.
Un ejemplo típico de este tipo de
bomba es la ATPasa de Na+-K+ o
Bomba de Na+-K+, responsable de
bombear Na+ fuera de la célula y K+
al interior de la misma y por lo tanto,
establece un pronunciado gradiente de
estos iones a través de la membrana
plasmática, acoplado a la hidrólisis de
ATP (Fig. 5).
Figura 5. Bomba de Na+-K+gghttp://1.bp.blogspot.com/_1e0dQzMNoaY/S58Vy-Csy/AAAAAAAABDs/SB9j_L_kxmw/s400/bomba+sodio+potasio.jpg
Para mantener el gradiente de
iones, las células gastan energía
considerable. Por ejemplo, más de
25% del ATP producido por las células
nerviosas y del riñón se utiliza para el
transporte de iones.
La bomba de Na+-K+ juega un
papel importante en la propagación de
señales eléctricas en el nervio y el
músculo. Además, el gradiente de Na+
establecido en la bomba, también se
emplea para dirigir el transporte activo
de otras moléculas como es el caso del
transporte de glucosa por las células
epiteliales intestinales. En la mayoría
de las células animales mantiene el
equilibrio osmótico y el volumen
celular.
Potencial de membrana o de
reposo, depolarizacion y potencial de
acción. Como los iones están cargados
eléctricamente, su transporte supone
que se establezca un gradiente o
potencial eléctrico a través de la
membrana. La magnitud del potencial
eléctrico varía entre -15 y -100 mV.
Para células no excitables este voltaje
se llama potencial de membrana y para
las células excitables como las

nerviosas y musculares, se denomina
potencial de reposo.
El potencial eléctrico se debe a
las bombas iónicas y al flujo de los
iones a través de los canales de la
membrana plasmática de la célula en
reposo. La membrana plasmática de la
célula en reposo, contiene canales de
K+ abiertos por lo que es más
permeable al K+ que al Na+ o a otros
iones. Como consecuencia, el flujo de
K+ supone la principal aportación al
potencial de membrana en reposo. La
concentración 20 veces superior en el
interior celular, respecto al fluido
extracelular, dirige el flujo de K+ hacia
el exterior celular. Como el K+ se
encuentra cargado positivamente el
flujo de este ión desde la célula, genera
un potencial eléctrico a través de la
membrana encontrándose el interior de
la misma cargada negativamente (Fig.
6).
Figura 6. Potencial de reposo.
http://recursos.cnice.mec.es/biosfera/alumno/1bachillerato/animal/imagenes/nervio/potenciacion.gif
Cuando se estimula la membrana
en reposo, la célula responde abriendo
la compuerta de algunos canales de
sodio y permite que penetre a la célula
un número limitado de iones sodio.
Este movimiento de cargas positivas al
interior celular reduce el potencial de
membrana que se vuelve menos
negativo. Al reducirse el voltaje
disminuye la polaridad entre los lados
de la membrana, lo que se denomina
despolarización. Luego que se abren
las compuertas del Na+ se cierran en
cuestión de milisegundos y se abren
los canales de K+, lo que provoca la
salida de K+ al exterior y el
restablecimiento del potencial de
reposo (Fig. 7). Los gradientes iónicos
se mantienen por la bomba Na+-K+
que utiliza la energía derivada de la
hidrólisis de ATP para transportar Na+
y K+ contra sus gradientes
electroquímicos.

Figura 7. Despolarización y repolarización http://www.mailxmail.com/curso-electrocardiografia-basica/anatomia-propiedades-celula-cardiaca-repolarizacion-despolarizacion
Los cambios en el potencial de
membrana luego de la despolarización
constituyen un potencial de acción .
La despolarización de las regiones
adyacentes de la membrana plasmática
permite a los potenciales de acción
viajar a lo largo de los axones de las
células nerviosas como señales
eléctricas, dando como resultado la
rápida transmisión de los impulsos
nerviosos a través de largas distancias.
Transporte activo secundario.
En este tipo de transporte se produce el
paso de sustancias a través de la
membrana celular que no son
permeables a la misma. Para lograr
esto, utilizan la energía del ATP de
manera indirecta. El ejemplo más
típico de este tipo de transporte es el
sistema de cotransporte sodio-glucosa
en el intestino delgado.
Cada molécula de glucosa que
se transporta desde el lumen o cavidad
del intestino delgado hacia el interior
del enterocito o célula intestinal, está
acompañada por el movimiento
simultáneo de un ión sodio. El paso de
este ión sodio se debe a que la bomba
Na+-K+ mantiene un gradiente de Na+
favorable. Es decir, la glucosa puede
transportarse en contra de un gradiente
de concentración a expensa del
transporte de sodio a favor de un
gradiente de concentración.
Endocitosis. Además de los
tipos de transporte descritos
anteriormente, las células eucariotas
también son capaces de captar
macromoléculas y partículas del medio
extracelular dentro de vesículas
derivadas de pliegues o invaginaciones
de la membrana plasmática. La
captación de materiales extracelulares
en vesículas citoplasmáticas se
denomina endocitosis. Cuando la
célula captura partículas grandes como
las bacterias, la endocitosis se
denomina fagocitosis (Fig. 8). Por otra

parte, la captación selectiva de
macromoléculas extracelulares
específicas (ligandos) luego de unirse a
receptores en la superficie exterior de
la membrana celular, se denomina
endocitosis mediada por receptores
(Fig. 9). Un ejemplo de este tipo de
transporte, lo constituye la captura de
colesterol por las células de mamíferos
que se transporta a través del torrente
sanguíneo en forma de lipoproteínas de
baja densidad (LDL). Para ello, se
requiere de de la unión de LDL a un
receptor específico y liberación
posterior del colesterol en la célula.
Figura8 . Fagocitosishttp://3.bp.blogspot.com/_bXiAT6MOo8E/S2syqWIgtlI/AAAAAAAACQE/C12uyaRa3mw/s400/fago01.png
Figura 9. Endocitosis mediada por un receptor.http://edu.jccm.es/ies/alonsoquijano/PaginaVieja/
websdelosdepartamentos/webdebiologiaygeologia/
imagenes/imagenes_biologia/MEMRECEP.GIF

ACTIVIDAD CONTROL 1
1. Identifica en el siguiente esquema, los componentes estructurales de la membrana
señalados con los números de 1 a 7.
2. Explique los factores que influyen la fluidez de la membrana
3. Elabora un esquema de llave que contemple los tipos de transporte de acuerdo al
gradiente de concentración y de acuerdo a la vía utilizada para atravesar la
membrana.
4. Analiza la función de las proteínas de membrana en el transporte de compuestos.
5. Describe el potencial de membrana en reposo, la depolarización y el potencial de
acción.
6. Analiza la importancia de la bomba Na+-K+.
7. Diferencia entre los procesos de fagocitosis y endocitosis mediada por receptores.

BIOENERGETICA Y METABOLISMO. MITOCONDRIAS
Las organelas que se encuentran
en el citoplasma celular no solo
cumplen funciones relacionadas con el
transporte y distribución de proteínas,
sino que además son compartimientos
especializados donde se llevan a cabo
diferentes actividades metabólicas.
Una de las actividades fundamentales
de todas las células es generar energía
metabólica, en forma de ATP. En las
células eucariotas animales las
mitocondrias son las organelas
responsables de generar la mayor
cantidad de energía, que es
almacenada en forma de ATP y
producida a partir de la degradación de
lípidos y carbohidratos. En las células
vegetales los cloroplastos utilizan la
energía solar para sintetizar
carbohidratos a partir de CO2 y H2O.
En células procariotas, la producción
de ATP se realiza en la membrana
plasmática.
GlucólisisLa glucólisis es el conjunto de
reacciones bioquímicas por medio de
las cuales la glucosa es metabolizada
hasta producir piruvato (compuesto de
tres átomos de carbono),
produciéndose energía metabólica
(Figura 10). En los tejidos animales, la
glucosa se transforma en piruvato, el
cual en presencia de oxígeno es
oxidado completamente en las
mitocondrias hasta CO2 y H2O. En éste
caso se habla de glucólisis aeróbica.
En ausencia de oxígeno, el piruvato
generado a partir de la glucosa es
transformado en lactato, en el proceso
denominado glucólisis anaerobia o
fermentación láctica.

Figura 10. Glucólisis: http://2.bp.blogspot.com/_HLVXQOUw3zs/TCQreC_7MAI/AAAAAAAAAH8/SUDMjU_6mxw/s1600/Glucolisis.png
La glucólisis es una de las rutas
catabólicas conocidas bajo el nombre
de fermentaciones anaeróbicas. Esta
secuencia de 10 reacciones
enzimáticas, es probablemente una de
las rutas bioquímicas mejor
comprendida. La glucólisis cumple una
función clave en el metabolismo
energético porque provee una cantidad
significativa de energía que es
utilizada por la mayoría de los
organismos y, por preparar a la
glucosa, así como a otros
carbohidratos, para la degradación
oxidativa.
Por medio de la glucólisis o
glicólisis, una gran diversidad de
organismos obtienen energía química a
partir de varios combustibles orgánicos
en ausencia de oxígeno molecular. La
mayoría de los organismos superiores
han conservado la capacidad de
efectuar la degradación anaeróbica de
la glucosa a lactato, que es una etapa
de preparación del catabolismo
aeróbico de la glucosa

En las células animales la
glucólisis desempeña un rol como
mecanismo de emergencia para la
producción de energía durante
períodos cortos en los que no se
dispone de oxígeno. Los organismos
heterótrofos (organismos que deben
alimentarse de sustancias orgánicas
sintetizadas por otros organismos)
obtienen su energía fundamentalmente
de las reacciones de oxido-reducción,
éstas son reacciones en las que los
electrones son transferidos desde un
compuesto, el dador de electrones o
agente reductor; a otro compuesto,
aceptor electrónico o agente oxidante.
Los organismos heterótrofos
aeróbicos obtienen la mayor parte de
la energía por medio de la
respiración, que es definida como la
oxidación de los combustibles
orgánicos por el oxígeno molecular; en
tal sentido, el oxígeno actúa como el
aceptor final de los electrones en la
respiración (Fig. 11).
Figura 11. Respiración celular.http://www.fisicanet.com.ar/biologia/metabolismo/ap1/oxidacion_glucosa01.jpg
Los organismos heterótrofos
anaeróbicos también obtienen la
mayor parte de su energía de las
reacciones de oxido-reducción, pero en
el proceso de fermentación. Los
electrones pasan desde un
intermediario orgánico producido
durante la degradación del azúcar, el
dador electrónico, hasta otro
intermediario orgánico quien es el
aceptor electrónico.
En la fermentación anaeróbica,
la D-glucosa es uno de los
combustibles más utilizados, sin
embargo, también se obtiene energía
de la fermentación anaeróbica de
ácidos grasos, aminoácidos, purinas, y
pirimidinas.
En la fermentación de la glucosa
predominan dos tipos que se
encuentran íntimamente vinculados: la

fermentación homoláctica y la
fermentación alcohólica.
En la fermentación
homoláctica la glucosa que es una
molécula de seis átomos de carbono es
degradada a dos moléculas de ácido
láctico (molécula de tres átomos de
carbono). Este tipo de fermentación se
produce en muchos microorganismos,
en la mayoría de los animales
superiores y en las plantas.
La fermentación alcohólica es
característica de muchas levaduras.
Durante éste proceso la molécula de
glucosa es escindida en dos moléculas
de etanol (cada molécula contiene dos
átomos de carbono), y dos moléculas
de CO2. En la fermentación alcohólica
ocurren las mismas reacciones que en
la glucólisis, pero precisa de dos etapas
enzimáticas diferentes al final de la
ruta. En su mayoría los demás tipos de
fermentación de la glucosa son
variaciones de la ruta fundamental de
la glucólisis.
La glucólisis es catalizada por la
acción consecutiva de 11 enzimas.
Estas enzimas pueden extraerse con
facilidad de las células en forma
soluble, por ésta razón se cree que las
enzimas glucolíticas se localizan en la
porción soluble del citoplasma. Al
parecer, las enzimas individuales que
catalizan las etapas de la glucólisis no
dependen físicamente unas de las
otras, en otras palabras, no se
encuentran formando complejos
multienzimáticos. Sin embargo, en
diferentes tipos celulares algunas
enzimas glucolíticas pueden
encontrarse unidas débilmente a la
membrana plasmática, a las
miofibrillas o a las mitocondrias.
En la glucólisis, todos los
intermediarios entre la glucosa y el
piruvato son compuestos fosforilados.
Estos grupos fosfato desempeñan tres
funciones:
1. Proveen a los intermediarios
glucolíticos un grupo polar con
carga negativa, lo que impide
que ésta molécula atraviese la
membrana. En efecto, a pH 7
los intermediarios de la
glucólisis son compuestos
iónicos, lo que impide que
atraviesen la membrana por
difusión simple.
2. Actúan como grupos de
reconocimiento para la

formación de los complejos
enzima-sustrato.
3. Permiten conservar la energía.
Al terminar la glucólisis esos
grupos fosfato se convierten en
el grupo fosfato terminal del
ATP.
En la glucólisis anaeróbica
primero se prepara la glucosa para su
catabolismo mediante su fosforilación,
posteriormente es escindida y forma
gliceraldehído-3-fosfato (molécula de
3 átomos de carbono). En una segunda
fase el gliceraldehído-3-fosfato se
convierte en lactato.
Durante la glucólisis ocurren tres
transformaciones químicas diferentes e
interconectadas:
1. La secuencia de reacciones a
través de las que el esqueleto
carbonado de glucosa es
degradado a lactato: la ruta de
los átomos de carbono.
2. La secuencia de reacciones
mediante las que el fosfato
inorgánico se transforma en el
fosfato terminal del ATP: la ruta
del fosfato.
3. La secuencia de óxido-
reducciones: la ruta de los
electrones.
Ciclo del Acido Cítrico
La mayor cantidad de ATP que
se forma durante el metabolismo de los
organismos superiores procede de la
fosforilación oxidativa, que se nutre
del potencial reductor o electrones, que
se liberan en la oxidación de distintos
sustratos a nivel de las membranas
intramitocondriales. Este proceso se
efectúa mediante la formación de
grupos acetilo, que en forma de acetil-
CoA (CH3-CO-SCoA o acetato activo)
son oxidados a CO2 y H2O mediante
una serie de reacciones conocidas
como Ciclo del Ácido Cítrico, ciclo de
los Ácidos Tricarboxílicos o Ciclo de
Krebs, que ocurre en las mitocondrias
(Fig. 12). Los electrones que son
liberados por la oxidación de los
grupos acetilo en el transcurso del
ciclo están disponible directamente
para la fosforilación oxidativa, lo que
permite la formación de hasta dos
terceras partes de todo el ATP que
consume el organismo.

En general el metabolismo se
divide en tres etapas. En la primera
etapa las macromoléculas tales como
las proteínas, polisacáridos y lípidos
son transformados en sus metabolitos
fundamentales: aminoácidos, glucosa,
glicerol y ácidos grasos. En la segunda
esos metabolitos son transformados
bien directamente o, previo a la
formación del piruvato en acetil-CoA.
Finalmente en la tercera etapa del
metabolismo, los dos átomos de
carbono del acetato activo entran en el
ciclo del ácido cítrico para su completa
oxidación a CO2, con la formación del
potencial reductor que nutre la
fosforilación oxidativa para la síntesis
de ATP y agua.
El Ciclo del Ácido Cítrico tiene
además una función anabólica, puesto
que es fuente de sustratos para la
síntesis de ácidos grasos, aminoácidos,
glucosa e incluso de la hemoglobina y
los ácidos nucleicos. También cumple
un papel importante en la regulación
metabólica de enzimas claves del
metabolismo.

Figura 12. Ciclo de Krebs.http://www.esacademic.com/pictures/eswiki/52/400px-Citric_acid_cycle_with_aconitate_2-es.svg.png
Cadena respiratoria o transportadora de electrones
Los organismos superiores
obtienen la energía de las sustancias
químicas sintetizadas por otros seres
vivos. La transferencia de esa energía a
sustancias químicas propias plantea
diversos desafíos. En principio, no solo
hay que aprovechar la mayor cantidad
de energía recibida sino que además
hay que almacenarla y dosificarla en
diversas formas. En tal sentido, la
molécula utilizada en la naturaleza
para tal fin es el ATP. Esto significa
que en el ATP se almacena la energía
proveniente de los diferentes nutrientes
y a partir del ATP la energía es
distribuida a los diferentes procesos
endergónicos de síntesis.
Para transferir la energía
proveniente de los distintos nutrientes
al ATP, en los seres vivos ocurre una
oxidación de los mismos. En tal
sentido, si la energía de
oxidorreducción se transfiere
directamente del sustrato que porta la
energía o nutriente al ATP, el
fenómeno se denomina fosforilación
unida al sustrato. En cambio, si la
energía de oxidorreducción en primer
lugar es acumulada en intermediarios
reducidos tales como las coenzimas
NADH(H+) y FADH2 o, raramente en
el NADPH(H+), y posteriormente, la
energía se utiliza para la síntesis de
ATP, el fenómeno se conoce como
fosforilación oxidativa, puesto que el
oxígeno es utilizado como último
aceptor de los electrones.
Termodinámicamente, la
fosforilación oxidativa ocurre debido
al acoplamiento de una reacción
fuertemente exergónica, es decir, la
transferencia de electrones desde las
coenzimas reducidas (NADH, FADH2
o NADPH) al oxígeno; a otra reacción
de carácter endergónico, es decir, la
síntesis de ATP. Es importante tener
presente que la cantidad de energía
liberada en las reacciones exergónicas
tiene que ser discreta para que pueda
ocurrir el acoplamiento energético. El
metabolismo celular solamente posee
la síntesis de ATP como la única

reacción endergónica capaz de
acoplarse directamente a los procesos
exergónicos. Dicho de otra forma, la
energía debe ser dispensada muy
cuidadosamente en paquetes
homólogos al enlace fosfato-fosfato
del ATP. A este respecto, si la energía
es liberada en paquetes de dimensiones
inadecuadas, no podría ser utilizada en
el trabajo químico y se disiparía en
forma de calor. Además, la liberación
de la energía debe estar sincronizada
con la síntesis del ATP, puesto que, la
energía aun cuando esté reducida a las
dimensiones adecuadas no encontraría
su destino. Para la dosificación de la
energía redox los organismos
aeróbicos poseen la cadena
respiratoria; y, para el acoplamiento de
la energía liberada con la síntesis de
ATP se utiliza la fosforilación
oxidativa (Figuras 13 y 14).
En los organismos aerobios la
cadena respiratoria y la fosforilación
oxidativa se localizan en la
mitocondria junto a la mayoría de las
deshidrogenasas. Esto significa que la
mitocondria está especializada en la
oxidación terminal de los sustratos.
Efectivamente, el carbono y el
hidrógeno, que forman
fundamentalmente los sustratos,
después del metabolismo mitocondrial,
se encuentran en la forma más
oxidada, es decir en CO2 y H2O. Este
hecho hace de las mitocondrias, las
centrales energéticas celulares, donde
se extrae la energía de los sustratos
energéticos, reduciéndolos a sustancias
de fácil eliminación.

Figura 13. Cadena transportadora de electrones y fosforilación oxidativahttp://recursos.cnice.mec.es/biosfera/alumno/2bachillerato/Fisiologia_celular/imagenes/cadenarespiratoria.gif
Figura 14. ATP sintetasa: formación de ATPhttp://4.bp.blogspot.com/_fXR9v67H4f8/S6_fNsBEzLI/AAAAAAAAACA/JIdBJAtx9Zs/s1600/ATP.jpg
MITOCONDRIAS
Como ya ha sido mencionado, en
las células eucariotas las mitocondrias
desempeñan un papel fundamental en
el proceso de generación de energía
metabólica. La mayor parte de la
energía que utiliza la célula proviene
de la degradación de los ácidos grasos
y de los carbohidratos que se realiza en
las mitocondrias, energía que es
convertida y almacenada en forma de

ATP durante el proceso de
fosforilación oxidativa.
Las mitocondrias están
delimitadas por un sistema de doble
membrana, la membrana mitocondrial
externa y la interna; situándose entre
ambas membranas el espacio
intermembrana. La membrana
mitocondrial interna posee numerosos
pliegues denominados crestas
mitocondriales que se proyectan hacia
la matriz mitocondrial, que es el
interior de la organela.
En la matriz mitocondrial se
encuentran las enzimas que catalizan el
metabolismo oxidativo, así como el
sistema genético mitocondrial. En las
células animales la fuente principal de
energía metabólica es la degradación
oxidativa de la glucosa y de los ácidos
grasos (Fig. 15).
Figura 15. Mitocondriahttp://4.bp.blogspot.com/_EdiSPJX1jg8/Sg4OQvq1jGI/AAAAAAAABtM/HUddNTjjKUA/s400/06-RC-02.jpg
Las etapas iniciales de la
degradación de la glucosa (glucólisis)
tienen lugar en el citoplasma, donde la
glucosa es convertida en piruvato,
compuesto que posteriormente es
transportado hacia la matriz
mitocondrial, donde es completamente
oxidado hasta CO2, produciendo la
mayor cantidad de energía a partir de
la glucosa, quien es posteriormente
almacenada en moléculas de ATP. En
éste proceso inicialmente se oxida el
piruvato a acetil CoA, que es
metabolizada en el Ciclo del Ácido
Cítrico en la matriz mitocondrial. En
tal sentido, el Ciclo del Ácido Cítrico
cumple un papel central en la

degradación oxidativa de los
carbohidratos y de los ácidos grasos.
Durante la oxidación de la acetil
CoA ocurre la reducción del NAD+ y
FAD a NADH y FADH2
respectivamente. La importancia de
ésta ruta metabólica reside en que la
mayor parte de la energía generada por
el metabolismo oxidativo se produce
durante la fosforilación oxidativa, que
ocurre en la membrana mitocondrial
interna. Los electrones de alta energía
del NADH y FADH2 son transferidos
al oxígeno molecular a través de la
cadena transportadora de electrones.
La energía que se produce durante las
reacciones de transferencia de
electrones, es convertida en energía
potencial acumulada bajo la forma de
un gradiente de protones a través de la
membrana mitocondrial interna, es
decir, en el espacio intermembrana,
energía que es utilizada para dirigir la
síntesis de ATP. Es decir, la membrana
mitocondrial interna representa el
lugar principal de generación de ATP.
La estructura de la membrana
mitocondrial interna refleja la
complejidad de sus funciones por
múltiples razones: las crestas
mitocondriales son la manifestación
morfológica del aumento de su
superficie, la presencia
excepcionalmente elevada de proteínas
(alrededor de 70 %), el hecho de ser
impermeable a la mayoría de los iones
y moléculas. Esta última característica
de la membrana es de importancia
determinante para mantener el
gradiente de protones que dirige la
fosforilación oxidativa. La membrana
mitocondrial interna constituye una
barrera funcional al paso de moléculas
de tamaño pequeño, entre el citosol y
la matriz mitocondrial, lo que permite
mantener el gradiente de protones que
dirige la fosforilación oxidativa
Actividades de control 2

1. Realice un dibujo de una mitocondria y señale las diferentes estructuras que
la conforman.
2. Nombre los productos finales de la glucólisis aeróbica y anaeróbica.
3. Qué rutas metabólicas toman los productos de la glucólisis en función de la
presencia o ausencia de oxígeno.
4. Explique cómo se acoplan la cadena de transporte de electrones y la
fosforilación oxidativa.
5. Explique la interrelación existente entre la glucólisis, el Ciclo de los Ácidos
Tricarboxílicos y la cadena transportadora de electrones.
BIBLIOGRAFIA
Boyer, Rodney. (2000). Conceptos en Bioquímica. México: Internacional Thomson Editores.
Casanova Peñuela, María del P. (2006). Investiguemos II Biología. Caracas: Editorial Excelencia C. A.
Cooper, Geoffrey M. y Hausman Robert E. (2008). La célula. (4ª ed.). Madrid: Marbán libros S. L.
Herrera, Emilio (Comp.). (1991). Bioquímica. Aspectos estructurales y vías metabólicas. (2ª ed.). Madrid: Interamericana McGraw-Hill.
Nelson, David; Cox, Michael M.; Lehninger, Albert L. (2005). Lehninger. Principios de Bioquímica. (4ª ed.). Madrid: Omega.
Proverbio, Fulgencio y Marín, Reinaldo. (2002). Biología 8vo. Caracas: Santillana

Proverbio, Fulgencio y Marín, Reinaldo. (2008). Biología 9no. Caracas: Santillana.
Teixeira, José M. (2004). Ciencias Biológicas Ciclo Diversificado 2. Caracas: Librería Editorial Salesiana S. A.