Proyecto SIP: 20070200 “Expresión de factores de...
37
1 Proyecto SIP: 20070200 “Expresión de factores de virulencia de Helicobacter pylori y Haemophilus influenzae, detección de Mycoplasmas y Ureaplasmas por PCR y detección de cepas DT 104 de Salmonella spp.” Dentro del programa: Aplicación de técnicas convencionales y moleculares en la caracterización y diagnóstico de bacterias de interés médico. SIP 440-SIP 553 coordinado por la Dra. en C. Graciela Castro Escarpulli Nota: de acuerdo a los objetivos, se individualizaron los resultados según cada protocolo en general, de acuerdo con cada grupo bacteriano se presenta como sigue; aunque en las tesis individuales terminadas y presentadas, pueden verificar los resultados con mayor extensión. Resumen extenso. 1. Introducción 1.1 Características de Helicobacter pylori El primer aislamiento de Helicobacter pylori (antes conocido como Campylobacter pylori) se hizo en 1982 por Marshall y Warren; es un bacilo Gram negativo pleomórfico (2.5-4.0 μm largo y 0.5-1.0 μm de ancho) que se observa curvo o en forma de “ala de gaviota”, es móvil por la presencia de cuatro a seis flagelos polares envainados de 30 μm de largo y 2.5 μm de grueso; crece en condiciones microaerofílicas entre los 3 y 5 días a 37°C, las colonias en los medios de cultivo son translúcidas y miden de 1 a 2 mm de diámetro .En cultivos sólidos y en medios líquidos, desarrolla formas cocoides, en cultivos viejos que son metabolicamente activas, sin embargo no son cultivables in vitro . El genoma de H. pylori tiene 1.67 Mb que codifica para 1500 proteínas. El contenido de G + C es del 37.5%. Aproximadamente el 40% de los aislados de H. pylori contienen plásmidos de 23.3 Kb, estos plásmidos no contienen factores de virulencia reconocidos, además el genoma posee por lo menos dos copias de cada rRNA 16S y 23S Los humanos también pueden ser infectados accidentalmente con H. heilmannii que es una bacteria espiral que se encuentra en perros, gatos, cerdos y primates pero no en humanos. La prevalencia en humanos de H. heilmannii es aproximadamente 0.5% causando una gastritis leve en la mayoría de casos, pero también se ha encontrado en relación con casos de linfoma de tejido linfoide asociado a mucosas (MALT) 2002). Los medios de cultivo sólidos utilizados para el aislamiento son gelosa sangre en medio base de Casman, gelosa chocolate, Skirrow, en general medios enriquecidos. El medio de cultivo líquido es caldo Brucella suplementado con suero fetal bovino al 10% . 1.2 Epidemiología. El humano es el hospedero definitivo del microorganismo, la prevalencia de la colonización por H. pylori se relaciona con la edad y la localización geográfica, cambia de países desarrollados a países en vías de desarrollo, siendo en estos, su prevalencia del 70 a 80% H.
Transcript of Proyecto SIP: 20070200 “Expresión de factores de...

1
Proyecto SIP: 20070200 “Expresión de factores de virulencia de Helicobacter
pylori y Haemophilus influenzae, detección de Mycoplasmas y Ureaplasmas
por PCR y detección de cepas DT 104 de Salmonella spp.”
Dentro del programa: Aplicación de técnicas convencionales y moleculares en la
caracterización y diagnóstico de bacterias de interés médico. SIP 440-SIP 553
coordinado por la Dra. en C. Graciela Castro Escarpulli
Nota: de acuerdo a los objetivos, se individualizaron los resultados según cada protocolo en general, de acuerdo con cada grupo bacteriano se presenta como sigue; aunque en las tesis individuales terminadas y presentadas, pueden verificar los resultados con mayor extensión.
Resumen extenso.
1. Introducción 1.1 Características de Helicobacter pylori El primer aislamiento de Helicobacter pylori (antes conocido como Campylobacter pylori) se hizo en 1982 por Marshall y Warren; es un bacilo Gram negativo pleomórfico (2.5-4.0 μm largo y 0.5-1.0 μm de ancho) que se observa curvo o en forma de “ala de gaviota”, es móvil por la presencia de cuatro a seis flagelos polares envainados de 30 µm de largo y 2.5 µm de grueso; crece en condiciones microaerofílicas entre los 3 y 5 días a 37°C, las colonias en los medios de cultivo son translúcidas y miden de 1 a 2 mm de diámetro .En cultivos sólidos y en medios líquidos, desarrolla formas cocoides, en cultivos viejos que son metabolicamente activas, sin embargo no son cultivables in vitro . El genoma de H. pylori tiene 1.67 Mb que codifica para 1500 proteínas. El contenido de G + C es del 37.5%. Aproximadamente el 40% de los aislados de H. pylori contienen plásmidos de 23.3 Kb, estos plásmidos no contienen factores de virulencia reconocidos, además el genoma posee por lo menos dos copias de cada rRNA 16S y 23S
Los humanos también pueden ser infectados accidentalmente con H. heilmannii que es una bacteria espiral que se encuentra en perros, gatos, cerdos y primates pero no en humanos. La prevalencia en humanos de H. heilmannii es aproximadamente 0.5% causando una gastritis leve en la mayoría de casos, pero también se ha encontrado en relación con casos de linfoma de tejido linfoide asociado a mucosas (MALT) 2002).
Los medios de cultivo sólidos utilizados para el aislamiento son gelosa sangre en medio base de Casman, gelosa chocolate, Skirrow, en general medios enriquecidos. El medio de cultivo líquido es caldo Brucella suplementado con suero fetal bovino al 10% .
1.2 Epidemiología. El humano es el hospedero definitivo del microorganismo, la prevalencia de la colonización por H. pylori se relaciona con la edad y la localización geográfica, cambia de países desarrollados a países en vías de desarrollo, siendo en estos, su prevalencia del 70 a 80% H.

2
pylori se adquiere en la niñez, usualmente antes de los 10 años de edad las personas no tratadas contra la infección pueden tenerlo en forma asintomática durante toda la vida. Niños, miembros de familias infectadas y aquellos que residen bajo condiciones de vida con niveles elevados de hacinamiento o que comparten cama con dos o más hermanos; además de una pobre higiene tienden a incrementar el riesgo de infección por H. pylori. Otros niños que tienen riesgo de infección elevada incluyen a aquellos de bajo nivel socioeconómico, niños nacidos de padres inmigrantes y niños adoptados internacionalmente provenientes de países pobres.
La incidencia de esta infección es mayor en afroamericanos e hispanos que en sujetos de raza blanca no hispanos, lo cual parece explicarse, solo en parte, por factores socioeconómicos. La prevalencia de H. pylori en el mundo (fig. 1) varia entre el 20% al 80%, con menor prevalencia de la infección en el Norte de América (Estados Unidos y Canadá) y Oeste de Europa, no obstante hay una prevalencia elevada en el Este de Europa, Asia países en vías de desarrollo y en muchos desarrollo como en México donde se tiene una prevalencia del 40 al 70% edades que van de en 5 - 9 años.
La infección es endémica en áreas con incidencia elevada de cáncer gástrico y esta asociado con casos de gastritis atrófica, que dan lugar a lesiones precursoras precancerosas con metaplasia que determinan más tarde el cáncer gástrico. En áreas del mundo con una baja frecuencia de cáncer gástrico, esta asociada con gastritis atrófica levey casos de úlcera gástrica o úlcera duodenal.
La Organización Mundial de la Salud, en 1994 clasificó a H. pylori como carcinógeno de tipoI para humanos y la agencia internacional de búsqueda de cáncer, consideró que el 43% de la carga global de cáncer gástrico pudiera estar relacionado con H. pylori .
Fig. 1. Prevalencia de la infección de Helicobacter pylori relacionado con el status socioeconómico en el mundo (URL-2)

3
1.3 Fuentes de infección.
La fuente de infección no se conoce con exactitud se plantea que es de humano-humano, por contaminación de comida y fuentes de agua de bebida o por vectores como la mosca (fig. 2). Se ha aislado de placa dental y vómito; H. pylori, puede sobrevivir hasta una semana en aguas residuales no potables y se ha relacionado la infección al consumo de vegetales crudos, regados con esta agua .
1.4 Rutas de transmisión
Las rutas de transmisión de H. pylori incluyen la fecal-oral, oral-oral y gástrica-oral. La identificación de H. pylori viable en vómito de adultos infectados, sugiere la posibilidad de que se produzca transmisión por vía gastrointestinal-oral durante episodios de gastroenteritis.
1.5 Patologías
H. pylori coloniza la mucosa gástrica son: capacidad microaerofílica, su forma, una espiral con flagelos polares envainados que le dan movilidad dentro de la capa de mucina y la actividad de la ureasa genera iones amonio proporcionando un ambiente alcalino que neutraliza el pH ácido del estómago. La historia natural de H. pylori puede evolucionar a partir de una gastritis superficial crónica e ir acompañada de dolor abdominal recurrente agudo (DAR). La infección produce gastritis antral (55%), úlcera duodenal (95%), úlceras gástricas (75%) y en casos graves carcinoma. Las lesiones por H. pylori representan como característica histológica una gastritis crónica activa superficial con infiltrado inflamatorio que afecta el epitelio secretor del moco y el cuerpo secretor del ácido, disminuyendo las defensas elementales de la mucosa .
Más del 90% de los pacientes con úlcera duodenal son portadores de H. pylori. La úlcera duodenal en ausencia de aspirina o fármacos antiinflamatorios no esteroideos (AINE) por lo
Fig. 2. Fuentes de infección de Helicobacter pylori (Frenck et al., 2003)

4
general se asocian con este microorganismo; puede colonizar el duodeno, pero solo se encuentra en islotes metaplásicos de epitelio del tipo gástrico (metaplasia gástrica). La colonización por H. pylori y la metaplasia gástrica se asocia con duodenitis activa, es una lesión precursora de la úlcera.
Las úlceras gástricas son benignas respecto de la úlcera duodenal que esta colonizada por H. pylori en una proporción menor de pacientes, la mayoría de las úlceras gástricas se atribuyen a la utilización de AINE o aspirina.
El linfoma tipo MALT es poco común tiene relación con la infección si se identifica pronto, en más de 75% , el tumor se puede curar con tratamiento eficaz contra la bacteria.
1.6 Factores de virulencia
Citotoxina vacuolizante (VacA): Aproximadamente el 50% de las cepas de H. pylori producen esta toxina. Esta citotoxina de 95kD se secreta como exotoxina, se inserta dentro la membrana celular y forma un canal selectivo de anión hexamérico que funciona como un tubo dependiente del voltaje, a través del cual son liberados aniones orgánicos, bicarbonato y así se obtiene el abastecimiento de nutrientes bacterianos, induce la formación de vacuolas ácidas en el citoplasma de células eucarióticas, que pueden ser identificadas al teñirse, porque acumulan rojo neutro, esto facilita su observación y análisis. La membrana mitocondrial se afecta por VacA y produce la liberación del citocromo “C” el que a su vez induce apoptosis.
El gen vacA codifica para una protoxina de 139-kDa contiene una secuencia señal de 33 aminoácidos tiene tres alelos diferentes (s1a, s1b, y s2), de la citotoxina (VacA) y un fragmento C-terminal de 50kDa con similitud al fragmento C-Terminal de la IgA precursora de Neisseria gonorrhoeae.
La región-m presenta a menos de 58 pares, varias secuencias “m” que están divididas en dos, m1 y m2. Las cepas s1/m1 son frecuentes y secretan altos niveles de la proteína VacA y son altamente toxigénicas en ensayos estandarizados de vacuolización con células HeLa.
Muchas cepas expresan el tipo s2 que falla al liberar la citotoxina. Las cepas que expresan s1/m2 producen cantidades significativas de citotoxina pero ésta no es tóxica para las células HeLa. H. pylori tiene el gen vacA, dan actividad de citotoxina pero en conjunto con las cepas que contienen la isla de patogenicidad (CagA). Se ha observado que mutantes VacA- pueden colonizar en modelos animales y cepas con el gen vacA inactivo pueden ser aisladas de pacientes, indicando que la proteína VacA no es esencial para la colonización.
Estudios sugieren que pacientes con cepas productoras de esta citotoxina desarrollan más frecuentemente úlcera péptica y adenocarcinoma gástrico.
Isla de patogenicidad (cag-PAI): Es un locus adquirido de 40kb denominado CagA, que está presente en el 60% de las cepas de H. pylori y se asocia con la expresión de la actividad citotóxica. CagA contiene 31 genes, varios codifican para componentes de un sistema de secreción tipo IV; que trasloca a la proteína CagA de 120kDa al interior de la célula huésped, afecta el citoesqueleto. Después de fijarse a las células epiteliales del estómago, la proteína CagA se inyecta dentro de la célula .La proteína se fosforila en un residuo específico de tirosina por proteínas de membrana de la célula huésped. El CagA traslocado tiene la

5
capacidad de inducir la producción de interleucina 8 (IL-8), que actúa dando lugar al reclutamiento y la activación de neutrófilos.
La presencia de CagA se asocia con alteraciones, como gastritis grave, atrofia de la mucosa y al riesgo de úlcera o cáncer gástrico. Las cepas procedentes de casos con úlcera, son cagA positivas en un porcentaje superior al 90%. Movilidad: La movilidad de H. pylori es un factor esencial para la colonización, normalmente esta bacteria posee de cuatro a seis flagelos polares envainados, cuyos filamentos consisten en dos tipos y están codificados por los genes flaA y flaB. Estos genes han sido clonados, induciéndoles mutaciones que han demostrado que ambos son esenciales para la completa movilidad.
Ureasa: La ureasa de H. pylori tiene aproximadamente 540kDa y esta formada por un hexamero que consiste en dos subunidades (UreA [30kDa] y UreB [62kDA]). La ureasa esta asociada con su membrana externa. La actividad de ureasa se necesita para la producción de un microambiente neutro adecuado dentro del lumen gástrico. H. pylori coloniza el estómago y la enzima ureasa hidroliza la urea a dióxido de carbono y amonio se convierte a bicarbonato que favorece y permite que la bacteria sobreviva a los efectos del ácido gástrico. La reacción de hidrólisis de la urea se da en la forma siguiente:
urea + ácido estomacal + agua --> bicarbonato + amonio
C=O·2 NH2 + H+ + 2H2O --> HCO3- + 2NH4
+
Catalasa y superoxido dismutasa: Los genes que codifican para la enzima catalasa y superóxido dismutasa de H. pylori tienen similitud con los genes de microorganismos intracelulares patógenos están en la superficie de H. pylori, estas enzimas protegen al microorganismo contra los efectos oxidantes del ión superóxido y del peróxido de hidrógeno endógeno, sugiriendo que tiene capacidad para exhibir una resistencia a la muerte mediada por leucocitos polimorfonucleares. 1.7 Helicobacter pylori en niños.
La barrera de ácido estomacal constituye una defensa contra patógenos entéricos, la destrucción de esta barrera puede hacer que los niños sean mas vulnerables a enfermedades diarreicas en países en desarrollo.
La asociación entre la colonización gástrica con H. pylori y la gastritis crónica en niños se ha confirmado en varios estudios y la úlcera gástrica y duodenal son raras. Los síntomas que aparecen con mayor frecuencia son dolor epigástrico denominado DAR dolor agudo recurrente, que constituye el motivo de consulta médica habitual, algunas veces va acompañado de vómito en aproximadamente la tercera parte de los niños. La prevalencia de factores de virulencia, como CagA y VacA, esta relacionada con la aparición de úlcera y cáncer gástrico en adultos, es menor en niños y aumenta proporcionalmente con la edad, lo que explica la menor incidencia de úlcera péptica en la población pediátrica .

6
Las infecciones tempranas en la niñez inducen un bajo grado de inflamación, esta condición se desarrolla poco a poco y produce cambios premalignos y eventualmente llega en algunos caso a desarrollar carcinoma gástrico.
La infección se relaciona con talla baja y retraso puberal en niñas preadolescentes, sin que hasta el momento se hayan podido demostrar cuales son los mecanismos implicados en estos casos. En contraste en la edad adulta cuando la infección se adquiere después de la niñez, es probable que se induzca una respuesta inflamatoria severa y aparezca una úlcera gástrica o duodenal .
1.8 Diagnóstico de la infección de Helicobacter pylori.
Hay dos tipos de pruebas de diagnóstico usadas para detectar la infección por H. pylori: técnicas invasivas y no invasivas. Las pruebas no invasivas detectan la presencia o ausencia de infección en forma indirecta, dentro de éstas se incluyen la prueba de aliento UBT (urea breath test), la determinación de anticuerpos en suero, y la prueba de antígeno en heces.
La prueba de aliento detecta CO2 marcado, producido por la hidrólisis de la urea, se usa en adultos como agente diagnóstico y para la confirmación de eliminación de la bacteria por el tratamiento, ya que la urea es un substrato importante necesario en el metabolismo de H. pylori que produce ureasa.
La realización del estudio involucra que el paciente tome una solución de urea marcada con 13C o 14C, en niños puede ser sustituido por zumo de naranja natural, seguida de la toma de la urea marcada. El átomo de carbono marcado (cualquiera de los dos 13C o 14C) son exhalados por el aliento como CO2 y se mide con aparatos especiales como el contador de centelleo o espectrofotómetro de masas. El 13C marcado es el más usado en niños, por ser un isótopo natural que no es tan radiactivo y puede emplearse sin riesgo de efectos secundarios; sin embargo, el marcaje y el aparato para detectarlos son muy caros .
Los métodos inmunológicos para la cuantificación de anticuerpos contra H. pylori en suero se basan en que la mayoría de los pacientes infectados manifiestan títulos elevados de anticuerpos IgG, IgA e IgM, dirigidos contra determinantes antigénicos de H. pylori. Se ha demostrado la presencia de IgA anti-H. pylori en jugo gástrico y saliva de los enfermos.
La prueba de antígeno en heces ha sido aprobada por la Food and Drug Administration (FDA) de EUA para evaluación de H. pylori. Su sensibilidad y especificidad es mayor del 90% en la detección de infección crónica; así como para confirmar la erradicación de la bacteria, cuatro semanas después de que se completa la terapia antibiótica.
Las pruebas invasoras determinan, la presencia o ausencia de la infección y la extensión y severidad de lesiones en la mucosa por métodos anatomopatológicos, dentro de éstas se incluyen la realización de endoscopia gastrointestinal con biopsia gástrica, a partir de ahí, se hace cultivo y se recupera a la bacteria H. pylori.
El estándar de oro para el diagnóstico de la infección de H. pylori es el cultivo del microorganismo a partir de las biopsias tomadas de la región antral del estómago, la positividad oscila entre el 80 y 90%.

7
1.9 Tratamiento. El tratamiento da tasas de erradicación superiores al 90%, puede ser de corta duración para asegurar el cumplimiento y con mínimos efectos secundarios aunque en general es prolongado.
La terapia triple con metronidazol, subsalicilato de bismuto o subcitrato de bismuto, y amoxicilina o tetraciclina durante 14 días, erradica la infección por H. pylori en 70 a 95% de los pacientes. La respuesta ha esta terapia se ve influida por su duración, en niños se han obtenido mejores resultados con pautas de dos semanas.
En adultos el tratamiento recomendado es la combinación de un antiácido con dos agentes antimicrobianos. En el caso de Francia, en donde las sales de bismuto no están autorizadas el tratamiento consiste en la asociación de la bomba inhibitoria de protones junto con amoxicilina o metronidazol. También se han observado resultados efectivos con inhibidores de la producción de ácido gástrico; ya que se provoca hiperácidez; cuando este esquema se completa con diferentes antibióticos como la claritromicina, amoxicilina, metronidazol.
El metronidazol es la base esencial del tratamiento, pero hay reportes internacionales que ponen de manifiesto resistencia, por lo que ha dejado de ser un antibiótico de primera elección, empleándose cada vez más la combinación de dos antibióticos: amoxicilina y claritromicina.
La respuesta al tratamiento debe ser monitorizada con la prueba del aliento con urea marcada con 13C, realizada entre cuatro y ocho semanas después de finalizar el tratamiento. En caso de fracaso terapéutico, será la presencia o no de sintomatología digestiva la que indique la instauración de una segunda pauta de tratamiento.
En España se emplea la terapia cuádruple con un inhibidor de la bomba de protones, bismuto, tetraciclina y metronidazol ante el fracaso de la terapia triple clásica, con una eficacia erradicadora media del 78%.
1.10 Mecanismos de resistencia antimicrobiana en la infección por Helicobacter pylori.
La infección bacteriana es un proceso complejo en el que tiene un papel importante, no solo la bacteria infecciosa, si no también el huésped; la respuesta del huésped a la misma altera ya que la homeostasis y la funcionalidad gástrica dando el cuadro clínico, algunos factores favorecen la infección; por ejemplo inmunodepresión, intubación, cateterización .
Las bacterias pueden ser resistentes a los antimicrobianos, o adquirir la resistencia bien por mutación, o como consecuencia de la adquisición de genes de resistencia que pueden residir en plásmidos que contengan otros genes que permitan se selección en ambientes en los cuales no haya presión selectiva por antimicrobianos.
Mecanismo de resitencia a macrólidos: Los macrólidos son un grupo de antibióticos con un anillo de lactona, la eritromicina es el macrólido más sencillo, son administrados oralmente difunden bien en el tejido, para el tratamiento de la infección por H. pylori en asociación con un segundo agente antimicrobiano, como amoxicilina o con nitroimidazoles y una droga antisecretora.

8
La claritromicina tiene un buen efecto in vitro, se afecta moderadamente por el decremento en el pH. Los macrólidos que han penetrado a la bacteria, se unen al ribosoma bloquea la síntesis de proteínas.
La claritromicina es un macrólido de 14 átomos de carbono tiene el anillo de lactona, en el carbono seis del anillo eritronólido hay la sustitución de un grupo metoxi que le da mayor estabilidad en el medio ácido, así como una mejor absorción y biodisponibilidad .
La claritromicina fue aprobada por la FDA en octubre de 1990, es activa contra Chlamydia pneumoniae y Legionella, así como, en Campylobacter spp y Helicobacter pylori.; es muy efectiva en las enfermedades de transmisión sexual y contra micobacterias atípicas. El mecanismo de acción de la claritromicina antibacteriana, interfiere en la síntesis de proteínas fijándose a la subunidad 50S ribosomal, da decremento de la afinidad de la claritromicina por los componentes del ribosoma 23S y por consiguiente hay una baja afinidad por el sitio blanco.
En el evento de la resistencia existe una disminución de la afinidad entre el ribosoma y la claritromicina, esta propiedad esta asociada con una mutación puntual en el gen 23s del rRNA en la posición 2143 y 2144. Esta mutación es esencialmente una transición de A-G pero puede haber una transversión de A-C en la posición 2143 en algunos casos.
Mecanismo de resistencia a nitroimidazoles: El metronidazol es uno de los representantes de este grupo, se absorben muy bien por vía oral y alcanza sus máximas concentraciones entre una y dos horas después de su administración oral; su vida media es de 8 h. Es un agente sintético antimicrobiano y antiparasitario, su indicación original fue para el tratamiento de infecciones provocadas por Trichomonas vaginalis; fue aprobado por la FDA para uso humano en 1963.
El metronidazol penetra en la bacteria; el grupo NO2 de los nitroimidazoles es reducido a la forma derivada de hidroxilamina. La reducción produce daño en el DNA causa la muerte de la bacteria.
El metronidazol tiene su acción antibacteriana por destructuración del DNA, la célula mediante difusión pasiva, baja el metabolismo anaerobio (proteínas de transporte de electrones de bajo potencial redox). El metronidazol reducido produce pérdida de la estructura helicoidal del DNA, ruptura de la cadena de inhibición de la síntesis de ácidos nucleicos y muerte celular.
Smith y Edwards en 1995 estudiaron H. pylori en presencia de metronidazol y encontraron que depende de la tensión relativa de oxígeno del medio ambiente y la densidad de la célula, determinan las condiciones redox del medio. Estos resultados concuerdan con los realizados por demostraron in vitro que la resistencia del metronidazol puede desaparecer cuando las células se incubadan anaeróbicamente a pocas horas del crecimiento. El desarrollo de la resistencia a metronidazol en cepas de H. pylori esta asociado por mutaciones (sin sentido), deleciones o inserciones en el gen rdxA, que codifica para una nitrorreductasa insensible al oxígeno que tiene la bacteria y que reduce el metronidazol, haciéndolo inactivo.

9
Resistencia a betalactámicos: La amoxicilina es el antimicrobiano más usado para la erradicación de H. pylori, fue aprobada por primera vez en 1981, es un antibiótico betalactámico, semisintético, que pertenece al subgrupo de las aminopenicilinas, es un derivado del núcleo alfa-amino-p-hidroxibencil penicilina con un radical amino (NH2
-) y un oxidrilo (OH-) en la posición del benceno. Este último, le permite una mejor absorción, distribución y concentración tanto en tejidos como en sangre.
Los antibióticos betalactámicos son agentes bactericidas que inhiben la síntesis de la pared celular bacteriana e inducen además un efecto autolítico. La destrucción de la pared celular bacteriana se produce como consecuencia de la inhibición de la última etapa de la síntesis del peptidoglucano, de este modo la pared queda debilitada y puede romperse por la presión osmótica intracelular. Al igual que todos los betalactámicos, la amoxicilina es susceptible de ser inactivada por enzimas (betalactamasas) que serán capaces de hidrolizar el anillo betalactámico y transformarlo en un anillo penicilinoico sin actividad antimicrobiana.
Para que la amoxicilina ejerza su mecanismo de acción, es fundamental que la bacteria se encuentre en pleno proceso de fisión binaria y la amoxicilina debe de tener integro su anillo betalactámico el cual, al pasar a través de las porinas de la bacteria, se fijará en la pared celular al inhibir las enzimas D-alanilcarboxipeptidasa y peptidoglican-transpeptidasa, impidiendo la formación de los enlaces cruzados en la síntesis de la pared celular bacteriana.
Una vez que el antibiótico betalactámico ha actuado, la falta de eslabonamiento de las peptidoglicana debilita la pared celular y forma un orificio provocando el entallamiento de la bacteria por la presión osmótica del exterior.
1.11 Antecedentes de resistencia y epidemiología de cepas resistentes en el mundo y en México.
El conocimiento de las enfermedades como gastritis crónica, úlcera péptica y posiblemente el cáncer gástrico fue acompañado por una evidencia clara relacionada con el fenómeno de resistencia a antibióticos, en donde la resistencia implica que antibióticos sin previa resistencia reportada por parte de la bacteria, ahora la presenten.
El tratamiento con antibióticos de H. pylori involucra la utilización de drogas usadas comúnmente para otro tipo de infecciones; por esta razón, el análisis del fenómeno de resistencia aparece desde la mitad del siglo pasado, coexiste con el uso de antibióticos y la bacteria que infecta, enfatizando la importancia del análisis de las variaciones geográficas y la evolución con el tiempo.
La resistencia de H. pylori a metronidazol y claritromicina; se ha incrementado a nivel mundial. La resistencia a amoxicilina ha sido reportada; sin embargo, este fenómeno es raro e inestable. La resistencia a amoxicilina desaparece después de un pase o después de la congelación prolongada de las cepas; en contraste, la resistencia a claritromicina y metronidazol queda igual después de varios pases o por congelación
Torres et. al durante el periodo de 1995 a 1997 en 195 niños y adultos de México, observo que la resistencia a metronidazol fue similar, en contraste la resistencia a amoxicilina y claritromicina se incremento significativamente durante este periodo de un 10% en 1995 a 27% en 1997, mientras que la resistencia a amoxicilina se incremento de un 13% en 1995 a 26% en 1997.

10
La resistencia a claritromicina se ha ido incrementando anualmente lo que sugiere que la prevalencia de la resistencia a claritromicina es más alta en cepas aisladas de niños, que de cepas aisladas de adultos; sin embargo, no se encontro diferencia entre estos grupos.
En México, el metronidazol usado comúnmente como un antiparásitario por muchos años, es una causa probable de la alta incidencia a la resistencia de este antibiótico. Otro estudio realizado de 1994 a 1999 con 150 niños de diferentes países, ahí la prevalencia de la resistencia a metronidazol y claritromicina antes del tratamiento a la infección con H. pylori fue elevada resistencia en niños originarios de África del 50% para metronidazol y en Europa del 29% para claritromicina.
Las cepas resistentes a claritromicina se han encontrado en niños menores de 10 años (10 al 19%). La resistencia a claritromicina en cepas de H. pylori provenientes de niños y adultos, sugiere la importancia del grupo de macrólidos en México y en Europa.
Susceptibilidad antimicrobiana y caracterización genotípica de aislados de Helicobacter pylori en el Hospital Infantil de México Federico Gómez. ALUMNA SANDRA MENDOZA
ELIZALDE D I R E C T O R E S : DRA. NORMA VELÁSQUEZ GUADARRAMA y DRA. SILVIA GIONO CEREZO
RESUMEN. El humano es el hospedero definitivo de Helicobacter pylori. la prevalencia de la colonización del microorganismo se relaciona con la edad y la localización geográfica, cambiando de países desarrollados a países en vías de desarrollo, por lo que H. pylori se adquiere en la niñez, antes de los 10 años de edad. Las personas no tratadas contra la infección pueden tenerlo en forma asintomática durante toda la vida. El tratamiento involucra antibióticos que en gran parte son usados comúnmente para otro tipo de infecciones. H. pylori puede ser tratado con un esquema que se completa con diferentes antibióticos como claritromicina, amoxicilina y metronidazol, los cuales fueron empleados para determinar la Concentración Inhibitoria Mínima. Resultados: Se trabajó con 86 cepas de H. pylori aisladas de pacientes pediátricos de HIMFG en el periodo del 2004 al 2007. Los resultados mostraron un 94.31% de resistencia para metronidazol y el 100% de sensibilidad para claritromicina y amoxicilina. Los genes de virulencia que posee H. pylori son los que le permiten colonizar a su hospedero mas fácilmente glmM, vacA, cagA, cagE; s1m1 cagA+ / cagE+ los cuales varían por región geográfica, por lo que se realizó la caracterización fenotípica de las cepas. El 72.6% (62/73) de las cepas fueron vacA s1m1 y presentaron un genotipo cagA+ / cagE+ el 75% ; (4/73) dieron un genotipo vacA s2m2 cagA+ / cagE+. Se encontraron cinco cepas con el genotipo vacA s2m1 cagA+ / cagE+, el cual no es muy común de forma natural. El riesgo de infección con múltiples poblaciones no está relacionado con la edad solamente, también está relacionado con varias características del hospedero, además de las condiciones ambientales que necesita la bacteria, tales como condiciones de vida o la higiene del hospedero. En este trabajo se pudo confirmar que un mismo paciente puede estar infectado por diferentes poblaciones, ya que la Concentración Inhibitoria Mínima dio valores diferentes, al igual que la caracterización de los genes de virulencia, lo que lleva a pensar que la erradicación del microorganismo es difícil debido a la diversidad de cepas, lo que facilita la selección de las mejor adaptadas al paciente y dificulta su erradicación.
PCR múltiple para marcadores de virulencia de Helicobacter pylori aisladas de pacientes pediátricos.

11
Introducción H. pylori es un bacilo Gram negativo, curvo y microaerofílico tiene morfología espiral en forma de sacacorchos. Mide de 0.5 a 1.0 μm de ancho y 3 μm de largo, tiene membrana externa. Produce ureasa, considerablemente más potente que en otras bacterias. Otras dos enzimas útiles para su identificación cuando crece en medios de cultivo son la oxidasa y la catalasa. H. pylori coloniza la mucosa gástrica del 50 % de la población mundial (Seok-Yong y cols., 2001) produce una infección crónica persistente. La infección por H. pylori se adquiere en la infancia desde el primer año. Las cepas se clasifican en dos grupos esta clasificación es útil al relacionar el genotipo de la cepa y la patogenicidad para establecer la relación con el curso clínico. Las cepas se han clasificado en: • Cepas tipo I: Cepas de H. pylori que poseen el gen cagA y expresan una citotoxina funcional (vacA). • Cepas tipo II: Cepas de H. pylori que perdieron el gen cagA y expresan una citotoxina (vacA) no funcional. Actualmente, se ha demostrado que la presencia genotípica de los marcadores: babA2, vacAs1 y cagA dan lugar a cepas triples-positivas, que muestran una elevada significancia estadística relacionada con la prevalencia de úlcera y adenocarcinoma. Marcadores de virulencia: Gen babA2: H. pylori que se adhiere a receptores al epitelio gástrico por medio de la adhesina BabA2 que es una adhesina de unión al antígeno de grupo sanguíneo Lewisb (_- 1,3/4-difucosilado) que se expresa en glóbulos rojos y en células del epitelio gástrico. El gen babA codifica para la adhesina, tiene dos alelos: babA1 y babA2. El gen que codifica para la adhesina completa y funcional es babA2; mientras que, el gen babA1 carece del codón de inicio. Gen cagA: Gen asociado a la citotoxina; se encuentra en el 50 al 60% de las cepas de H.pylori. El gen cagA, se emplea como marcador de la isla de patogenicidad (cagA-PAI) que codifica para un sistema de secreción tipo IV y el incremento en la secreción de IL-8. La infección de H.pylori con cepas cagA,+ aumenta el riesgo de úlcera duodenal y adenocarcinoma. Gen vacA: El gen vacA codifica para una citotoxina vacuolizante; sin embargo solo el 50% de las cepas expresan la citotoxina activa. Su diversidad involucra dos regiones: la región s con dos tipos alélicos: s1 y s2; a su vez, s1 posee los subtipos: s1a, s1b y s1c mientras que la región m tiene dos tipos alélicos: m1 y m2. Los alelos s1/m1 producen altos niveles de toxina; s1/m2 producen niveles bajos o moderados de toxina mientras que, las cepas s2/m2, no producen una toxina activa. La citotoxina, se ha aislado del 66.6% de pacientes con padecimientos como: úlcera péptica, frente al 30.1% presente solamente en el grupo con gastritis crónica. Métodos y materiales La población de estudio fueron 127 clonas aisladas de 18 biopsias de pacientes pediátricos pertenecientes a la colección del Laboratorio de Bacteriología Médica, proporcionadas por el Departamento de Endoscopia Pediátrica HGCM. “La Raza” del Instituto Mexicano de Seguro Social (IMSS) con la colaboración de la Dra. Rosalía Austria Míreles. Resultados Diseño del iniciador GLM MR1: El diseño del iniciador reverso para la detección del gen glmM se realizó con base en las secuencias reportadas en el “GeneBank” con número de acceso AE001446. Se realizaron

12
análisis en el programa DNAMAN para analizar las interacciones entre los iniciadores. Los iniciadores empleados para detectar el gen ureA al igual que para el gen glmM, no mostraron complementariedad. El iniciador se denominó GLM MR1 (5´- GCA TTC ACA AAC TTA TCC CCA ATC-3´) y amplifica un producto de 140 pb. Técnica de hibridación “Southern blot”: La capacidad de detección de las sondas fue: cagA (100 pg/μl), vacAm1 (100 pg/μl), vacAs1 (100 pg/μl) y babA2 (1 pg/μl). Los resultados demostraron que los amplicones correspondieron a los obtenidos por PCR. Secuenciación: El reporte escrito demostró que, el valor máximo de puntuación que fue de 1047, el valor E fue de 0 en la mayoría de las secuencias de los genes validados por lo que se puede considerar un alineamiento significativo. Los resultados mostraron que las secuencias amplificadas corresponden perfectamente a los productos babA2, cagA, vacAm1/s1, glmM de H. pylori; por lo que, la técnica de PCR múltiple se aplicó para caracterizar las cepas clínicas. Aplicación en las cepas clínicas: Los genes: cagA, vacAm1, vacAs1, se detectaron con mayor frecuencia, los datos se analizaron con el programa estadístico epiInfo™ 2000. El resultado fue: vacAs1 +(71.63 %), vacAm1+(68.5 %), cagA+ (66.14 %) semejante a lo observado en población adulta; el gen babA2+ se encontró en menor proporción (37.79 %). Los marcadores de virulencia: vacAm2/s2 se encontraron en menor porcentaje; sin embargo, se relacionaron con casos de UP. Las cepas tipificadas por PCR se clasificaron para establecer el porcentaje y frecuencia de cada genotipo en la población estudiada según la clasificación de Atherton y Gerhard: cepas tipo I: babA2-, cagA+, vacAm1s1+ (40.15%); cepas tipo II babA2-, cagA-, vacAm2s2 +(22%); triple positivas babA2+, cagA+, vacAm1s1+ (28.34%). En este estudio también se obtuvieron los genotipos: babA2+, cagA+, vacAs1+ (n= 4); babA2 +, cagA-, vacAm2s2+ (n= 8), estas cepas no se clasificaron por la ausencia o presencia de un marcador en particular. Las cepas del paciente B046: B046C1, B046C5, B046C9, B046C11, B046C12, B046C13, B046C14, B046C15 fueron cepas triple positivas con el genotipo: babA2 +, cagA+, vacAm1s1+ . 2. H. pylori ATCC 43504, 3. BO46C1, 4. B046C5, 5. BO46C9, 6. B046C11, 7. BO46C12, 8. B046C13, 9. B046C14, 10. B046C15. La distribución del gen babA2 en las cepas genotipificadas babA2 se detectó en 48 cepas y se distribuyó principalmente en triple positivas (n= 36) también, se detectó en las cepas: babA2+, cagA+, vacAs1+ (n= 4); cagA-, babA2+ vacAm2s2+(n= 8). Sensibilidad y especificidad del ensayo múltiple Se genotificaron 127 cepas H. pylori por PCR simple y múltiple el objetivo fue determinar la sensibilidad y especificidad del ensayo múltiple. La sensibilidad fue del 100% para todos los casos excepto para babA2 que fue de 31.25%; la especificidad fue del 100%. La PCR individual y PCR duplex sirvio para detectar los genes ureA y glmM, en las 104 muestras de DNA y en el DNA extraído de las 6 cepas recuperadas del cepario mostraron una alta sensibilidad y especificidad, no así cuando se aplico la PCR duplex Impacto: • Las cepas aisladas de la población infantil mexicana estudiada presentaron dos genotipos no clasificables: babA2
+, cagA+, vacAs1

13
+; babA2
+, cagA-, vacAm2s2
+. • La población infantil estudiada se encontró colonizada con mayor frecuencia con cepas tipo I; sin embargo, también se detectaron cepas triples positivas. Las cepas tipo II amplificaron el gen cagE por lo que, probablemente podría emplearse como mejor marcador de la isla cag-PAI que cagA. • El gen babA2 se encontró distribuido en cepas triple positivas y a diferencia de la bibliografía no se observó relación entre úlcera péptica y estas cepas; sin embargo, fueron aisladas de pacientes con padecimientos crónicos. • El iniciador diseñado GLM MR1 muestra alta especificidad para detectar el gen glmM , por lo que se puede utilizar para identificar a H. pylori. La técnica de PCR duplex puede ser empleada para la detección de H. pylori en muestras clínicas o en otro tipo de muestra. 2. Métodos y Materiales
o 86 cepas de Helicobacter pylori provenientes de aislamientos clínicos de biopsias gástricas obtenidas por endoscopia.
o Cepa de referencia Helicobacter pylori ATCC 43504 2.2. Diagrama general de trabajo
Biopsia Gástrica
Obtención de mezcla de poblaciones de las cepas
CIM de cepas de
H. pylori identificadas
Aislamiento de cepas puras de H. pylori
Claritromicina
Amoxicilina Caracterización genotípica glmM, cagA, cagE, vacA.
Extracción de DNA (kit)

14
2.3.1- Cultivo bacteriano a partir de biopsias gástricas Las muestras de biopsia gástrica y duodenales fueron colectadas en caldo Brucella suplementado con suero de caballo 10% y transportadas al laboratorio en un tiempo no mayor de 3 horas.
Las muestras fueron maceradas en un mortero de vidrio esmerilado, con caldo Brucella, el material el macerado se inoculó en placas de agar base de Casman suplementado con sangre de caballo 7.5% con y sin antibiótico para disminuir el sobrecrecimiento de cualquier flora competitiva. Las placas se incubaron a 37°C por 3- 5 días en atmósfera de 10%CO2. 2.3.2 Identificación bacteriana
La identificación bacteriana se realizó por morfología colonial, a partir de cajas de agar base de Casman con sangre de caballo al 7.5% y suplemento antimicrobiano. Las placas se incubaron en un ambiente microaerofílico de 3 a 5 días a 37° C; se seleccionaron colonias típicas circulares, elevadas, borde entero, grisáceas como “gotas de rocío”.
A partir de las colonias seleccionadas se realizó tinción de Gram, en donde se observaron los morfotipos de H. pylori como bacilo Gram negativo cuya forma característica es en “ala de gaviota” o en forma de “s”. Se realizó la prueba de ureasa observando el cambio de color del indicador rojo de fenol de ácido (amarillo) a alcalino (rosa mexicano) indicativo de la hidrólisis de la urea por la enzima ureasa de H. pylori. También se utilizaron las pruebas de catalasa y oxidasa que fueron positivas para la bacteria. 2.3.3.- Aislamiento de cepas Las cepas obtenidas se sembraron en agar base de Casman con sangre de caballo 7.5%, para obtener colonias aisladas, se incubo a 37º C por 3 días. Después de la incubación con ayuda de una asa bacteriológica se picaron 5 colonias aisladas de cada cepa, para posteriormente sembrarlas en una caja de agar base de Casman con sangre de caballo 7.5% que se incubo a 37º C por 3 días. La identificación numérica de las cepas se realizó dando un número progresivo del 1 al 5 (por ejemplo cepa 24: 24/1, 24/2, 24/3, 24/4, 24/5). 2.3.4.- Conservación de las cepas
De cada cepa aislada se sembró en masivo una caja de agar base de Casman, posteriormente se recolecto la biomasa para ser conservada en criotubos de 1.5 ml. Cada criotubo contenía 800 μl de caldo Brucella con suero fetal bovino al 10% y 25% de glicerol. Los criotubos se conservaron en congelación a -70°C (apéndice 9.3).
2.3.5.- Ensayos de susceptibilidad a los antimicrobianos
2.3.5.1.- Determinación de la Concentración Inhibitoria Mínima (CIM) por dilución seriada en placa de agar con replicador de “Steers” .
Se determinó la CIM para amoxicilina, metronidazol y claritromicina, empleando la dilución seriada en placas de agar con replicador de “Steers”, se empleó la cepa de referencia H. pylori ATCC 43504.

15
Las cepas se descongelaron y crecieron en cajas de agar base de Casman con sangre de caballo 7.5% en atmósfera de CO2 de 3 a 5 días a 37° C. Se cosechó el crecimiento en caldo Brucella o Mueller Hinton y se llevó a una suspensión bacteriana semejante al tubo 2.0 del nefelómetro de MacFarland con una concentración de bacterias 1x107 – 1x108 UFC/ ml. Las diluciones del antimicrobiano seleccionado se ajustaron en las placas de agar base de Mueller Hinton con 10% de sangre de caballo (apéndice 9.7) a las diferentes concentraciones. Para la replicación se ocupó 1000μl de la suspensión bacteriana previamente ajustada de cada cepa en los diferentes pozos del replicador “Steers” colocando cuidadosamente la parte superior del replicador (tapa) para poder llevar a cabo la replicación (por duplicado), se incubó las placas en atmósfera de CO2 de 3 a 5 días a 37° C.
2.3.5.2- Lectura de resultados y valores de corte. La CIM es la concentración mas baja del antibiótico donde se inhibe el crecimiento aparente del 99% de las bacterias. En la dilución seriada en placa con el replicador de “Steers”, la CIM está indicada por la placa con menor concentración de antibiótico que no presentó crecimiento de la cepa. Los valores de corte para considerar a las cepas sensibles o resistentes a los antimicrobianos fueron los siguientes:
• Amoxicilina: resistente ≥ 4 μg / ml (Torres et al., 2001). • Metronidazol: resistente ≥ 8 μg / ml (Torres et al., 2001). • Claritromicina: resistente ≥ 1 μg / ml (Kobayashi et al., 2001).
La cepa de referencia utilizada para el ensayo de susceptibilidad a los antimicrobianos fue:
• Cepa de referencia Helicobacter pylori ATCC 43504 Aislamiento: antro gástrico humano (Australia) Características genotípicas: cagA+, vacA+, s1am1. Susceptibilidad a antimicrobianos:
Amoxicilina sensible ( CIM 0.016 - 0.12 μg / ml )
Metronidazol resistente ( CIM 64 - 256 μg / ml )
Claritromicina sensible ( CIM 0.016 - 0.12 μg / ml )
2.4. Identificación de genes por técnicas moleculares en cepas de Helicobacter pylori
2.4.1.- Extracción de DNA genómico.
La extracción de DNA de H. pylori a partir de cultivo se realizó por el método comercial de Wizard Purificación Genómica del DNA (kit-Promega).
• Método comercial de Wizard Purificación genómica del DNA (kit). 1.- En un tubo eppendorf se adicionó 200 μl de suspensión de la cepa, 100μl de lisis nuclear y 100 μl de lisis celular, se incubó a 80°C por 5 minutos y se dejo enfriar.
2.- Se adicionó 3 μl RNAsa, que se dejó incubar por 15 minutos a 37°C.
3.- Se agregó 50 μl de solución de precipitación de proteínas (Vortex) y se dejó incubar 5 minutos en hielo.

16
4.- Se agregó 500 μl de cloroformo:alcohol isoamílico 24:1 (se agitó vigorosamente) para después centrifugar a 14000 rpm / 10 minutos.
5.- Se recuperó la fase acuosa y se transfirió a un tubo nuevo.
6.- Se adiciono 600 μl de isopropanol (frió) y se dejó toda la noche a –70°C.
7.- Se centrifugó a 14000 rpm por 5 minutos para decantar y adicionar Etanol al 70% (3 lavados).
8.- Se decanto el Etanol (último lavado) y se dejó secar al aire.
9.- Se rehidrató con 50 μl de agua inyectable.
2.4.2 Caracterización genotípica de aislamientos de Helicobacter pylori.
Se realizó por medio de la PCR, la amplificación de los genes de virulencia de
H. pylori : glmM, cagA , cagE (en este trabajo) y vacA a cada cepa aislada.
En la tabla 1 se muestran los iniciadores utilizados, condiciones y el producto esperado para la amplificación de los diferentes genes de virulencia. Posteriormente se realizó el corrimiento electroforético de los fragmentos de DNA amplificados en un gel de agarosa al 1.2%, el cual se tiñó con bromuro de etidio por 15 minutos en agitación constante.
Tabla 1. Condiciones de PCR
GEN OLIGONUCLEÓTIDOS CONDICIONES
DE LA PCR
NÚMERO
DE
CICLOS
PRODUCTO
glmM
glmM-F
5-AAGCTTTTAGGGGTGTTAGGGGTTT3
glmM-R
5-AAGCTTACTTTCTAACACTAACGC-3
(Smith et al, 2004)
93°C 45 seg
57°C 45 seg
72°C 45 seg
35 294 pb.
cagA cagA-s
5-AATACACCAACGCCTCCAAG-3
cagA-as
5-TTGTTGCCGCTTTTGCTCTC-3
(Lage et al, 1995)
94°C 1 min
55°C 1 min
72°C 1 min
35 400 pb.
cagE cagE-s
5-TGTGGCAAGCAAACAAGCT-3
94°C 45 seg 35 195 pb.
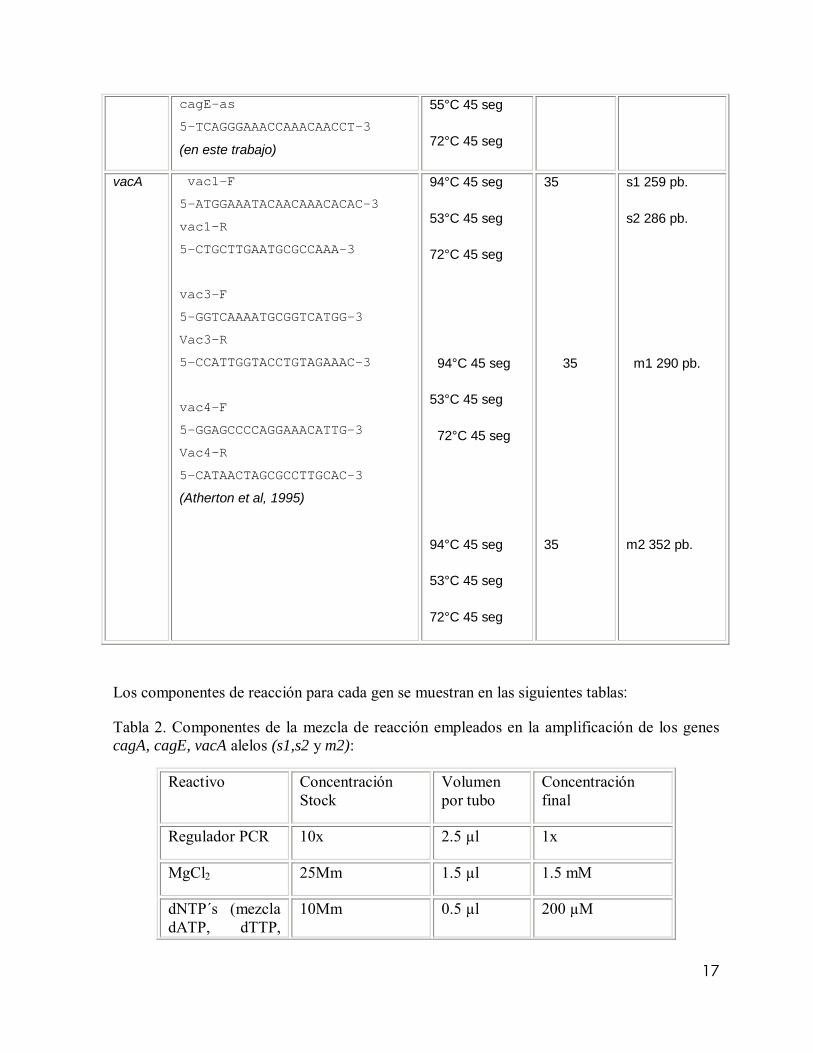
17
cagE-as
5-TCAGGGAAACCAAACAACCT-3
(en este trabajo)
55°C 45 seg
72°C 45 seg
vacA vac1-F
5-ATGGAAATACAACAAACACAC-3
vac1-R
5-CTGCTTGAATGCGCCAAA-3
vac3-F
5-GGTCAAAATGCGGTCATGG-3
Vac3-R
5-CCATTGGTACCTGTAGAAAC-3
vac4-F
5-GGAGCCCCAGGAAACATTG-3
Vac4-R
5-CATAACTAGCGCCTTGCAC-3
(Atherton et al, 1995)
94°C 45 seg
53°C 45 seg
72°C 45 seg
94°C 45 seg
53°C 45 seg
72°C 45 seg
94°C 45 seg
53°C 45 seg
72°C 45 seg
35
35
35
s1 259 pb.
s2 286 pb.
m1 290 pb.
m2 352 pb.
Los componentes de reacción para cada gen se muestran en las siguientes tablas:
Tabla 2. Componentes de la mezcla de reacción empleados en la amplificación de los genes cagA, cagE, vacA alelos (s1,s2 y m2):
Reactivo Concentración Stock
Volumen por tubo
Concentración final
Regulador PCR 10x 2.5 µl 1x
MgCl2 25Mm 1.5 µl 1.5 mM
dNTP´s (mezcla dATP, dTTP,
10Mm 0.5 µl 200 µM

18
dGTP, dCTP)
Iniciador
cagA-s
cagA-as
cagE-s
cagE-as
vac1-F
vac1-R
vac4-F
vac4-R
20 pmoles
20 pmoles
20 pmoles
20 pmoles
20 pmoles
20 pmoles
20 pmoles
20 pmoles
2 µl
2 µl
2 µl
2 µl
2 µl
2 µl
2 µl
2 µl
0.8 pmoles/ µl
0.8 pmoles/ µl
0.8 pmoles/ µl
0.8 pmoles/ µl
0.8 pmoles/ µl
0.8 pmoles/ µl
0.8 pmoles/ µl
0.8 pmoles/ µl
Taq DNA polimerasa
5 U/ µl 0.3 µl 1.5 U
H2O inyectable c.b.p 25 µl 14.2 µl -------------
DNA genómico 200 ng/ µl 5 µl 40 ng
Tabla 3. Componentes de la mezcla de reacción empleados en la amplificación de los genes glmM y vacA alelo (m1):
Reactivo Concentración Stock
Volumen por tubo
Concentración final
Regulador PCR 10x 2.5 µl 1x
MgCl2 25Mm 1.5 µl 1.5 mM
dNTP´s (mezcla dATP, dTTP, Dgtp, dCTP)
10mM 0.5 µl 200 µM
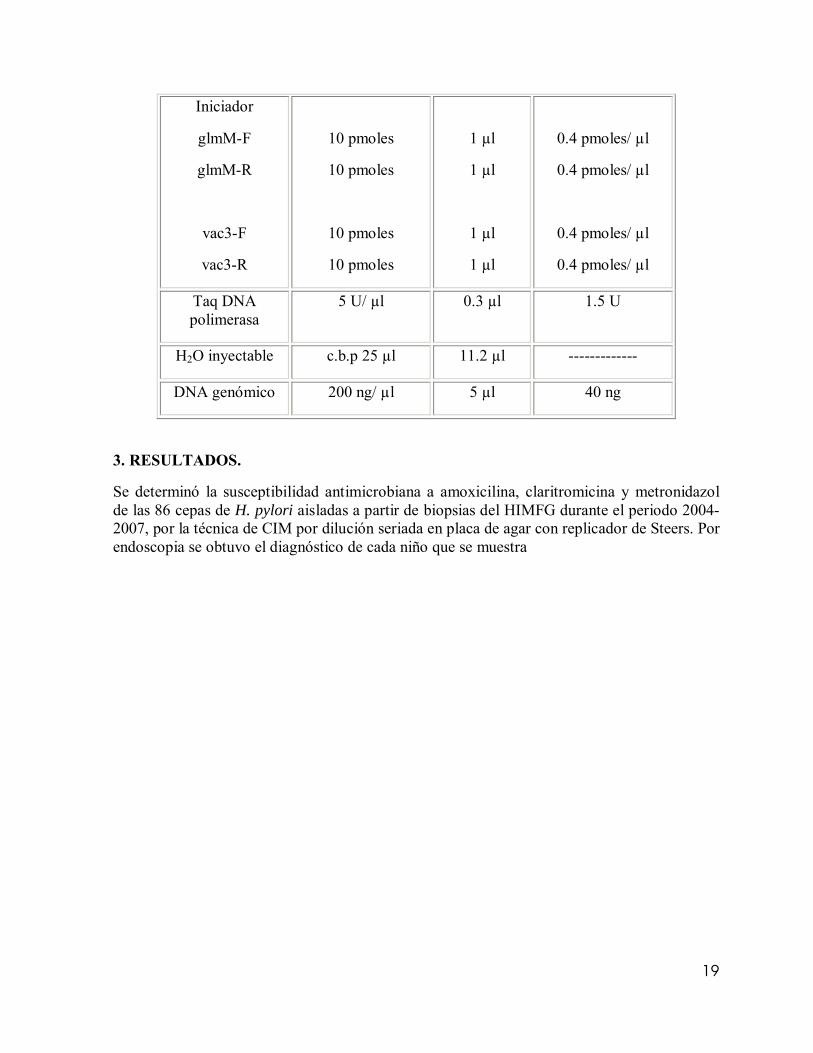
19
Iniciador
glmM-F
glmM-R
vac3-F
vac3-R
10 pmoles
10 pmoles
10 pmoles
10 pmoles
1 µl
1 µl
1 µl
1 µl
0.4 pmoles/ µl
0.4 pmoles/ µl
0.4 pmoles/ µl
0.4 pmoles/ µl
Taq DNA polimerasa
5 U/ µl 0.3 µl 1.5 U
H2O inyectable c.b.p 25 µl 11.2 µl -------------
DNA genómico 200 ng/ µl 5 µl 40 ng
3. RESULTADOS.
Se determinó la susceptibilidad antimicrobiana a amoxicilina, claritromicina y metronidazol de las 86 cepas de H. pylori aisladas a partir de biopsias del HIMFG durante el periodo 2004-2007, por la técnica de CIM por dilución seriada en placa de agar con replicador de Steers. Por endoscopia se obtuvo el diagnóstico de cada niño que se muestra

20
En la figura 6 se muestra el porcentaje de sensibilidad obtenido de cada cepa de
H. pylori por la técnica de CMI, mostrando una sensibilidad del 100% para amoxicilina y claritromicina y un 5.68% para metronidazol.
100%
5,68%
100%
0%10%20%30%40%50%60%70%80%90%
100%
amoxicilina metronidazol claritromicina
En la tabla 5 se muestra el número de cepas que presentaron resistencia y sensibilidad a amoxicilina, claritromicina y metronidazol en la técnica de CIM.
Tabla 5. Cepas que presentaron resistencia y sensibilidad por CIM a los tres antibióticos empleados.
Antibiotico No. cepas
resistentes
No. cepas
sensibles
Total
Amoxicilina 0 86 86
Metronidazol 81 5 86
Claritromicina 0 86 86
En la figura 7 se muestra la distribución de la CMI de los tres antibióticos empleados para las 86 cepas de H. pylori aisladas del HIMFG durante el periodo 2004-2007.
Fig. 6. Porcentaje de sensibilidad de cepas de Helicobacter pylori

21
0
10
20
30
40
50
60
70
0,01
0,02
0,03
0,06
0,13
0,25 0,
5 1 2 4 8 16 32 64 128
256
512
CIM
No. d
e ce
pas
Amoxicilina Metronidazol Claritromicina
Tabla 7. Frecuencia de genotipos glmM, vacA, cagA y cagE en 86 cepas de
Helicobacter pylori.
GEN ALELO NÚMERO (%)
glmM 86 100%
Total 86
s1 71 84.52%
s2 13 15.47%
Total 84*
m1 69 92%
vacA
m2 6 8%
Fig. 7. Distribución de la CIM de 86 cepas de Helicobacter pylori aisladas del HIMFG del 2004-2007

22
Total 75*
cagA positivo 79 91.86%
cagA negativo 7 8.13%
Total 86
cagE positivo 64 74.41%
cagE negativo 22 25.58%
Cag PAI
Total 86
vacA* 2 cepas no fueron tipificables para los alelos s1, s2; 11 cepas no fueron tipificables para los alelos m1,m2. En la tabla 8 se muestra la frecuencia de combinaciones alélicas, encontradas para los genes vacA, cagA y cagE en las cepas de H. pylori aisladas de biopsias del HIMFG.
Tabla 8. Frecuencia de las diferentes combinaciones alélicas encontradas para los genes vacA, cagA y cagE en las cepas de Helicobacter pylori.
Gen vacA cagA+ /cagE+
n (%)
cagA-/ cagE-
n (%)
cagA+/ cagE-
n (%)
cagA-/ cagE+
n (%)
Total (%)
s1m1 45 (72.58) 0 (0) 16 (25.80) 1 (1.6) 62
s2m2 3 (75) 0 (0) 1 (25) 0 (0) 4
s1m2 0 (0) 1 (50) 1 (50) 0 (0) 2
s2m1 5 (100) 0 (0) 0 (0) 0 (0) 5
A continuación se muestran algunos electroferogramas realizados para cada gen de las cepas de Helicobacter pylori:
100 pb.
200 pb.
294 pb.

23
Fig. 8. Electroferograma de los productos de PCR del gen glmM de cepas de Helicobacter pylori. carril: (1) marcador de peso molecular (100pb.); (2) Control negativo; (3) cepa 24/1; (4) ATCC 43504; (5) cepa 60/1; (6) cepa 20A/1; (7) cepa 10/1; (8) cepa 74/1; (9) cepa 20A/5 ; (10) cepa 24/3.
Fig. 9. Electroferograma de los productos de PCR del gen cagA de cepas de Helicobacter pylori. carril: (1) marcador de peso molecular (100pb.); (2) cepa 80/1; (3) cepa 24/2; (4) cepa 7/2; (5) cepa 4/2; (6) 5/3; (7) cepa 24/3; (8) cepa 32/1; (9) cepa 4/4; (10) cepa 99/5.
1 2 3 4 5 6 7 8 9
500 pb.
400 pb 400 pb.

24
Fig. 10. Electroferograma de los productos de PCR del gen cagE de cepas de Helicobacter pylori. carril: (1) marcador de peso molecular (100pb.); (2) cepa 74/5; (3) cepa 80/4; (4) cepa 80/3; (5) cepa 74/4; (6) cepa 1/1; (7) cepa 1/2; (8) cepa 5/1; (9) cepa 5/3; (10) cepa 5/4; (11) marcador de peso molecular (100pb.); (12) cepa 37/5; (13) cepa 79/1; (14) cepa 79/2; (15) cepa 4/3; (16) cepa 4/4; (17) cepa 4/5; (18) cepa 77/3; (19) cepa 77/1; (20) cepa 77/2.
1 2 3 4 5 6 7 8 9
11 12 13 14 15 16 17 18 19
300 pb.
200 pb. 195 pb.
195 pb. 300 pb.
200 pb

25
Fig. 11. Electroferograma de los productos de PCR del alelo s1, s2 de cepas de Helicobacter pylori. carril: (1) marcador de peso molecular (100pb.); (2) cepa 24/1; (3) cepa 24/2; (4) cepa 24/3; (5) cepa 24/4; (6) cepa 37/4; (7) cepa 2/4; (8) cepa 37/5; (9) cepa 37/1; (10) cepa 37/2; (11) cepa 37/3; (12) cepa 1/1; (13) cepa 1/2; (14) cepa 10/1; (15) cepa 10/2; (16) cepa 10/3; (17) cepa 60/1; (18) cepa 60/2; (19) cepa 60/3; (20) cepa 77/1.
Fig. 12. Electroferograma de los productos de PCR del alelo m1 de cepas de Helicobacter pylori. carril: (1) marcador de peso molecular (100pb.); (2) cepa 24/3; (3) cepa 37/4; (4) cepa 24/2; (5) cepa 37/1.
4. Discusión. En este trabajo se evaluó la Concentración Inhibitoria Mínima de tres antibióticos usados en la terapia triple de H. pylori en pacientes pediátricos del Hospital Infantil de México Federico Gómez, ocupando 86 cepas las cuales arrojaron resultados de resistencia de más del 70% para metronidazol y sensibilidad del 100% para amoxicilina y claritromicina. Se evaluaron los genes de virulencia que le confieren una mayor capacidad de adaptación al microorganismo y el posible desarrollo de úlcera péptica y/o cáncer gástrico, mostrando una mayor prevalencia del genotipo vacA s1/m1 cagA positivo/ cagE positivo. Se estudiaron 86 cepas de Helicobacter pylori aisladas de biopsias de niños pacientes del Hospital Infantil de México Federico Gómez del periodo 2004-2007, debido a que es un microorganismo que se adquiere en la edad temprana (antes de los 10 años de edad) La
1 2 3 4 5
290 pb.
400 pb.
300 pb.

26
mayoría de los pacientes al diagnóstico presentó enfermedad ácido péptica, dos presentaron hiporexia, uno enfermedad por reflujo esofágico y uno ambas enfermedades.
A las 86 cepas se les determinó la Concentración Inhibitoria Mínima (CIM) a diferentes antibióticos por dilución seriada en placa de agar con replicador de “Steers” (CLSI 2005). Los antibióticos utilizados fueron metronidazol (nitroimidazol), amoxicilina (betalactamico) y claritromicina (macrólido); ya que estos son los frecuentemente utilizados como terapia combinada para la erradicación de H. pylori.
Los resultados de CIM (tabla 4) de las 86 cepas fue para metronidazol, del 94.31% resistentes y 5.69% sensibles, para claritromicina y amoxicilina el 100% de las 86 fueron sensibles. En metronidazol existen diversos estudios alrededor del mundo en donde se ha observado que más del 50% de las cepas aisladas de H. pylori son resistentes . En el caso de México, ha aumentado hasta en un 70% su resistencia, datos que son corroborados por los obtenidos en este estudio, que mostraron que las cepas probadas presentan resistencia en el 94.31%. Esto puede deberse a que el metronidazol es utilizado como tratamiento antiparasitario, lo que llevaría a ocasionar la resistencia del microorganismo por el uso frecuente del mismo. La amoxicilina la gran mayoría de las cepas son sensibles pero la resistencia pudiera perderse por continuos pases o por descongelmiento de las cepas. Los resultados mostraron 100% de sensibilidad hacia este antibiótico, pudiéndose presentar este fenómeno, ya que las cepas estuvieron en continuo descongelamiemto para el uso de este protocolo.
En el caso de claritromicina al igual que metronidazol se ha visto en los últimos años, que la resistencia de las cepas de Helicobacter pylori han ido en aumento. Sin embargo, en nuestro caso todos los aislamientos fueron sensibles a claritromicina observando dos eventos distintos: 1) pacientes multitratados o que han presentado fallas en el tratamiento y que continúan con un diagnóstico positivo lo que hace difícil el aislamiento del microorganismo in vitro, y 2) la gran mayoría de los pacientes que se les ha logrado aislar al microorganismo no han recibido tratamiento para H. pylori, en la gran mayoría de ellos logran la erradicación del microorganismo si hay apego al mismo.
En la caracterización de los genes de virulencia de las cepas de H. pylori, se encontró una alta frecuencia de cepas cagA positivo (91%) y cagE positivo (74%) (tabla 7) y con esto hubo una asociación importante entre estos dos genes, lo que sugiere que la mayoría de los aislados de la población pediátrica tienen una isla cag-PAI completa y que es potencialmente funcional. Se encontró también una mayor prevalencia de cepas con genotipo vacA s1m1 (tabla 5). Se determinó la frecuencia de asociación entre los alelos s1 y m1 y los alelos s2 y m2 del gen vacA (tabla 8) arrojando resultados del 84.93% (62/73) de cepas que presentaron genotipo s1/m1, el 5.47% (4/73) genotipo s2/m2, el 2.73% (2/73) genotipo s1/m2 y el 6.84% (5/73) vacA s2/m1.
En la tabla 8 se muestra la frecuencia de las diferentes combinaciones alélicas encontradas para los genes vacA, cagA y cagE, observándose una asociación importante entre los genes cagA y cagE y el genotipo vacA s1m1 pues el 72.58% (62/73) de las cepas vacAs1m1 presentó un genotipo cagA positivo/ cagE positivo y el 75% (4/73) de los aislamientos

27
vacA s2m2 tuvieron un genotipo cagA positivo/ cagE positivo. En el caso de los genotipos heterologos vacA s1m2 cagA positivo/ cagE negativo se presentó el 50% (1/2), y para vacA s2m1 cagA positivo/ cagE positivo el 100% (5/5).
Al igual que estudios anteriores, la presencia de la isla de patogenicidad cag se correlacionó con una mayor actividad citotóxica por el genotipo vacA s1/m1 cagA positivo/ cagE positivo 72.58%, que esto nos refleja una producción de la citotoxina vacA funcional y la presencia de cag-PAI, que son caracteristicas asociadas con una mayor virulencia de H. Pylori, se encontraron cinco cepas con el genotipo vacA s2m1 cagA positivo/ cagE positivo, el cual no es muy común. Sin embargo en el estudio hecho por Morales-Espinosa et al., 1999 se reportó la presencia de este genotipo, el cual fué corroborado en este estudio.
El riesgo de infección con múltiples poblaciones no está relacionado solo con la edad también intervienen varias características del hospedero y condiciones ambientales.
En este trabajo se confirmó que un mismo paciente puede estar infectado por diferentes poblaciones, ya que la CIM arrojó valores diferentes para amoxicilina, claritromicina y metronidazol, al igual que en la caracterización de los genes de virulencia, lo que nos lleva a pensar, que esta sea la razón de lo difícil que es la erradicación del microorganismo.
β-lactamasas Tipo TEM1 en cepas de Haemophilus influenzae Objetivo - Comparar la eficacia del medio HTM contra el medio de GC durante las pruebas de susceptibilidad por CMI en placa.- Identificar las especies y serotipos - Identificar el modelo de resistencia de Hi a antibióticos β-lactámicos con Difusión de Disco y CMI- Identificar el gen blaTEM1 que codifica a la enzima β-lactamasa TEM1 y la presencia del plásmido p1056 Materiales y Métodos Se elaboraron 4 lotes de placas de HTM y de GC para CMI de ampicilina de acuerdo a las recomendaciones del CLSI 2005. Un par de lotes (GC y HTM) fue probado al siguiente día de ser preparado, el otro par de lotes se uso 7 días después. Se utilizaron 23 cepas previamente caracterizadas por difusión de disco y Cefinasa®; 14 cepas fueron resistentes y hubo 9 sensibles Cefinasa® negativas; 13 cepas fueron Cefinasa® positivas y 1 negativa, Todas fueron inoculadas con cultivos de 18h. Se analizaron 115 cepas de distintas fuentes de aislamiento la tabla 1 muestra los orígenes y los serotipos. 75 cepas fueron aisladas en 2003-2004. 40 fueron aisladas en 1998. Se aglutinaron con antisueros: polivalente y contra serotipo b. La resistencia se hizo se hizo con difusión de disco y la CMI en dilución de placa. La producción de β-lactamasa con discos de nitrocefina Cefinasa®. Se realizó una PCR duplex para detectar al gen blaTEM1 y plásmido p1056. Los iniciadores usados fueron de Hasegawa et al 2003 y Leaves et al 2000. Los amplificados fueron visualizados por electroforesis de gel de agarosa por métodos convencionales. Los medios HTM Y GC, con las 23 cepas se desarrollaron bien en las categorías a las que corresponden. Las 14 cepas resistentes dieron en el rango de 8-128μg/ml y las 9 sensibles de 0.015-0.25μg/ml, las mismas cepas probadas con el medio de HTM, solo crecio el 65.2% del total de las cepas y el rango obtenido fue de 0.015-0.064μg/ml. Mientras que, en GC las 23 cepas se desarrollaron bien, 14 cepas resistentes dieron en el rango de 4-32μg/ml manteniéndose en la categoría de resistentes. Se obtuvo un 28.7% (33/115) de cepas resistentes en la población analizada. La población aislada en 2003-2004,mostró un 20.1% de cepas resistentes (24/75), 21.2% (9/40).

28
Se obtuvieron 24 cepas con el gen blaTEM1 y 19 con el plásmidos, solo 48.9% (16/33) mostraron tanto el plásmido como el gen blaTEM1. vio desplazada hacia la derecha, lo cual indica que las cepas no han disminuido su susceptibilidad. Hubo 2 cepas que mostraron un patrón de resistencia cefuroxime, cefotaxime y ceftriaxona; además de ampicilina, una de ellas no mostró blaTEM1 pero fue Cefinasa® positivo. Una cepa aislada de nasofaríngeo fue resistente a amoxicilina/ácido clavulánico y ampicilina pero fue Cefinasa® negativa. Se observó además que hubo una prevalencia del gen blaTEM1 en cepas capsuladas (18.2% Hib y 18.2% de capsuladas no b). En cepas NTHi hubo 6 cepas blaTEM1
negativas pero Cefinasa® positivas. 5 cepas que amplificaron el p1056 dieron amplificados de peso molecular (~500bp) más alto que el reportado (370bp). Impacto El medio de HTM es un medio que utiliza ingredientes puros como el β-NAD, este componente es muy inestable en soluciones acuosas, tal inestabilidad aumenta cuando se adiciona a medios con una temperatura mayor a 40º C. Si es almacenado en refrigeración es estable solo durante 2 semanas en soluciones no en medios. El medio de GC es una mezcla compleja de distintos componentes, sin embargo es mucho más estable que el NAD utilizado del medio HTM de modo que permite que los microorganismos se desarrollen bien aunque disminuya un poco la eficacia para promover el crecimiento, pero tal disminución no fue representativa, porque las cepas resistentes continuaron creciendo en la misma categoría. La presencia del gen blaTEM1 se asocia con la producción de β-lactamasas TEM1, fue la prevalencia mayor en los aislados de Hi, se observo que fue mayor en las capsuladas que en las NTHi. Solo hubo una cepa Cefinasa® negativa relacionada con un fenotipo low BLNAR porque demostró resistencia amoxicilina/ácido clavulánico resistente, pero no dio resistencia a las cefalosporinas.. El modelo de resistencia que prevaleció en este grupo fue el de resistencia por β-lactamasas de tipo TEM1 sobre las cepas Cefinasa® positivas blaTEM1
negativas. Las cepas NTHi verdaderas, es decir carecen de cápsula fueron genéticamente más variables, este fenómeno se observó en 6 cepas blaTEM1 negativas, 3 presentaron también el plásmido p1056. El plásmido presente en cepas del serotipo b; estuvo en 4 de enfermedad sistémica (2 de hemocultivo y 2 LCR). Los iniciadores que se utilizaron para la detección del p1056 abarcan desde un fago similar a CTX de V. cholerae hasta una región de repetidos invertidos, es posible que los amplificados de mayor peso molecular sean resultado de una duplicación de los repetidos invertidos o que el fago similar a CTX, tenga información complementaria que haga que el amplificado sea de un peso molecular más alto. Impacto - El medio de HTM es un medio útil si se utiliza hasta 24 después haberse preparado, de otra forma, el medio da resultados falsos que pueden aparecer como sensibles - El medio de GC demostró mantener el desarrollo de las 23 cepas probadas y dentro de sus respectivas categorías (14 cepas resistentes y 9 sensibles) - Es necesario realizar pruebas de control de calidad en los laboratorios en los que se utilice este medio. • La resistencia por β-lactamasas ha reducido hasta 21.3% en las cepas estudiadas de 2003-2004 comparado con el 48.5% de Giono et al. 2001. • El modelo de resistencia que prevalece es de cepas BLPAR de tipo TEM1. Es posible que las cepas con el fenotipo de resistente a CTX, CXM, CRO y AMP presenten una β-lactamasa distinta a TEM1 las descritas anteriormente. • La susceptibilidad a los β-lactámicos fue reducida, la moda de cepas susceptibles es de 0.031μg/ml y la CMI50% de 0.125 μg/ml.

29
• La presencia del plásmido es posible que se relacione con la presencia de cápsula, ya que en estas cepas no hay presencia del plásmido si este no esta acompañado del gen blaTEM1. • Las secuencias que amplificaron con un peso molecular de 500bp mayor al reportado es posible que contengan ya sea información complementaria del fago similar a CTX o que solo tengan duplicados de las secuencias repetidas directas. ADHERENCIA DE Haemophilus influenzae NO TIPIFICABLE (NTHi) DE ORIGEN CLÍNICO Y SU RELACIÓN CON EL GEN hap Introducción Haemophilus influenzae es una bacteria Gram negativa identificada como patógeno importante en población infantil y de adultos. La patología como agente etiológico, es clasificada como: enfermedades severas, agudas, piógenas, generalmente invasivas y enfermedades crónicas. Haemophilus influenzae posee mecanismos de adherencia, para penetrar y colonizar células del epitelio respiratorio humano. La superficie bacteriana se une a diferentes receptores de las células huésped durante la infección, resultando en una colonización por: ADHESINAS; entre ellas están: pili, IgA proteasa, proteína Hap, proteínas HMWI y HMW2 (proteínas de alto peso molecular), proteínas OPM´S (proteínas de membrana externa), proteína Hia, lipooligosacárido (LOS) y lipopolisacárido (LPS) La proteína Hap (Homologous IgA protease Protein) es una proteína autotransportadora perteneciente al sistema de secreción tipo V. Es una serin-proteasa identificada por su habilidad para promover una interacción íntima en cultivos de células epiteliales humanas. Se traslada como una proteína precursora de 155 kDa, con tres dominios: una típica secuencia iniciadora N-terminal, una fracción de 110 kDa serin- proteasa (nombrada HapS) y una proteína de membrana externa C-terminal de 45 kDa (denominada Hapβ). El gen hap es ubicuo para cepas de H. influenzae, y Hap es producida por todos los aislamientos no tipificables. OBJETIVO Determinar el patrón de adherencia de cepas NTHi en cepas de origen clínico mexicanas; y relacionarlo con la presencia del gen hap. Métodos y materiales. a) Cepas de referencia H. influenzae ATCC 49766, Hi ATCC 49247, Hi ATCC 39930, E.coli K12 y E. coli agregativa (proporcionadas en el lab. de Bacteriología Médica) b) 20 Cepas de Haemophilus influenzae no tipificables (NTHI) de origen clínico mexicanas (proporcionadas por el Hospital Infantil de México y la Benemérita Universidad de Puebla). De las cuales 9 son del serotipo b, 4 son capsulares no b y 7 son no tipificables (NTHi) c) Línea celular HEp- 2 (carcinoma epidermoide de laringe humana). In Vitro, S.A. ENSAYOS DE ADHERENCIA. A partir de una capa semiconfluente de células HEp-2 sembradas dentro de placas de 24 pozos se estandarizó la técnica de adherencia con cepas ATCC (H. influenzae 33930, 49247 y 49766), se ocupó como testigo positivo la cepa E. coli agregativa y como negativo la cepa E. coli K12. La bacteria se inoculó en un caldo y se incubó hasta obtener una densidad aproximadamente de 2x109 UFC/ml. Se inoculó 1x107 UFC en la monocapa de células epiteliales. Después de una incubación de 90 minutos a 37°C en 5% de CO2, las monocapas se lavaron 5 veces con PBS (pH 7) para remover las bacterias que no se adhirieron, se fijaron con metanol absoluto, se tiñeron con GIEMSA, se observaron al microscopio para contar las células con adherencia bacteriana por campo y se hizo el cálculo de porcentaje de adherencia (St. Geme, J.W. III y cols., 1993).

30
TÉCNICA DE AMPLIFICACIÓN EN CADENA DE LA POLIMERASA (PCR) Se utilizó la técnica de extracción de DNA de Tiocianato de Guanidina. Se estandarizó la técnica utilizando los iniciadores propuestos por Kilian y cols. (2002). Se determinó la presencia del gen hap en cepas de Haemophilus influenzae no tipificables (NTHi) de origen clínico. La presencia del gen hap se determinó en el 55% (11 de 20) de las cepas clínicas; 72.7% (8 de 11) fueron del serotipo b, 18.2% (2 de 11) fueron capsulares no b y 9.1% (1 de 11) fueron no tipificables (NTHi). Estudios previos han determinado que el gen hap esta presente en todas las cepas de H. influenzae; sin embargo, nuestros resultados determinaron ausencia del gen hap en sólo 1 de 9 cepas del serotipo b, en 2 de 4 cepas capsulares no b y en 6 de 7 cepas NTHi. Impacto La relación adherencia- Hap se ve reflejada con la cepa de referencia Hi ATCC 33930; que presentó mayor adherencia y fue positiva para hap. La relación vacuolización- Hap se observó también con la cepa Hi ATCC 33930; que presentó mayor vacuolización y fue positiva para hap. El daño celular puede estar relacionado con la autoproteólisis de Haps; lo que pudo afectar a la integridad de las células. Se ha planteado que la liberación de Hap facilita la degradación de células blanco del huésped, así como de componentes del tejido o induce expresión efectores del sistema inmune (Henderson y cols., 2004). En la PCR para la identificación de hap en cepas clínicas, se determinó que en las cepas del serotipo b Solo 1/9 resultó hap – La presencia del gen hap en 8 cepas del tipo b pudo considerarse como serotipos virulentos. Las cepas capsulares no b 2/4 (50%) y no tipificables 6/7 (85%) fueron negativas para el gen hap. La presencia de hap es un factor importante en la adherencia de Haemophilus influenzae en células HEp-2. La citotoxicidad (vacuolización) en células HEp-2 pudiera estar relacionada con la presencia de hap debido a la autoproteólisis de Haps El 55% de las cepas clínicas fueron hap positivas, predominando cepas del tipo b, lo que pudiera sugerir una relación entre el gen hap y cepas virulentas. La presencia del gen hap en 8 cepas del tipo b pudo considerarse como serotipos virulentos. Las cepas capsulares no b 2/4 (50%) y no tipificables 6/7 (85%) fueron negativas para el gen hap. La presencia del gen hap se determinó en el 55% (11 de 20) de las cepas clínicas; 72.7% (8 de 11) fueron del serotipo b, 18.2% (2 de 11) fueron capsulares no b y 9.1% (1 de 11) fueron no tipificables (NTHi). Estudios previos han determinado que el gen hap esta presente en todas las cepas de H. influenzae; sin embargo, nuestros resultados determinaron ausencia del gen hap en sólo 1 de 9 cepas del serotipo b, en 2 de 4 cepas capsulares no b y en 6 de 7 cepas NTHi. La relación adherencia- Hap se ve reflejada con la cepa de referencia Hi ATCC. Determinación de Salmonella Typhimurium DT 104 en México, patrones de resistencia antimicrobiana y PFGE Periodo Junio-Agosto 2006. RESUMEN El género Salmonella spp comprende más de 2,000 serovares distintos, responsables de salmonelosis en el hombre y animales. A través de los años ha sufrido cambios en su taxonomia, y en la actualidad se clasifica en Salmonella enterica y Salmonella bongori con sus diversos serovares. Es una de las bacterias patógenas que puede transmitirse por la ingestión de agua y alimentos contaminados, causando una amenaza a la salud. Es responsable de enfermedades como gastroenteritis, bacteremia y fiebre entérica (tifoidea y paratifoidea). Algunos serovares pueden infectar a las aves, cerdos, reptiles y becerros. En los últimos años se ha descubierto y caracterizado un gran

31
número de elementos genéticos de Salmonella spp conocidos como “Islas de Patogenicidad”, las cuales se encuentran agrupadas en zonas específicas del cromosoma bacteriano que codifica factores de virulencia. Se involucran en los procesos de entrada de la bacteria a células epiteliales y de sobrevivencia en el interior de los macrófagos del hospedero. Existe evidencia experimental que sugiere que estas regiones han sido adquiridas mediante transferencia horizontal del material genético. El aislamiento e identificación de Salmonella spp se realiza mediante métodos de cultivo tradicionales, serológicos (reacciones febriles, coaglutinación, ELISA, inmunofluorescencia) y métodos moleculares como hibridación con RNA y PCR. Las técnicas de ingeniería genética en el estudio de la patogenicidad bacteriana ha abierto un nuevo campo en el análisis de la virulencia a nivel molecular, este conocimiento es de gran valor para el desarrollo de nuevos medicamentos y vacunas. Debido a que la salmonelosis continúa representando un problema de salud pública en los países desarrollados y en vías de desarrollo, se hace imprescindible implementar métodos de identificación para la prevención y control de la enfermedad, así como estrategias que mejoren las condiciones sanitarias de la población
Muestras y procesos de rutina en casos sospechosos de salmonelosis. Hemocultivo.- Útil para el diagnóstico de fiebre tifoidea, no resulta difícil aislar a S. Typhi si se realiza durante la primera semana después del inicio de los síntomas y antes de iniciar el tratamiento, tiene un éxito del 90%. Se inócula de 3 a 5 mL de sangre sospechosa en medio Ruiz-Castañeda o medios adecuados para el aislamiento de enterobacterias y se incuba horizontalmente a 37 °C en aparatos automatizados como Bact Sept, Check etc., durante 21 días revisándose diariamente ya que por lo general se recupera en las primeras

32
24 a 72 h, si se desarrollan colonias sobre la fase sólida se aísla e identifican 8. 43 Cuadro Muestras y procesos de rutina en casos sospechosos de salmonelosis. Coprocultivo.- Cuando se sospecha de salmonelosis o fiebre tifoidea, la muestra debe ser tomada en la etapa aguda de la enfermedad y antes del tratamiento antimicrobiano. La materia fecal en estudio se siembra en medios de cultivo como agar Mac Conkey, agar Salmonella–Shigella y sulfito de bismuto, y en caldos de enriquecimiento como tetrationato o caldo selenito. Se incuba 18-24 h a 37 °C. Se resiembra en medios de alta selectividad como agar verde brillante y agar sulfito de bismuto, seleccionar las colonias sospechosas y se hacen pruebas bioquímicas, por resiembra en tubos con los medios MIO, LIA y TSI 8. Los resultados obtenidos en las bioquímicas se comparan con los cuadros de identificación de Salmonella spp. Una vez obtenida la cepa pura se procede a su serotipificación. Salmonella spp, al igual que otras enterobacterias poseen antígenos de superficie (H y Vi) que interfieren con la aglutinación por antisueros O y se relaciona con la invasividad de la bacteria. Los antígenos somáticos más frecuentemente aislados en México son los grupos B, C1, C2, D y E 8. 7.2 Diagnóstico Serológico. Confirmación serológica de dichas colonias. Serotipificación de Salmonella spp. Se prueban con suero polivalente de Salmonella A-I y después con los 5 antisueros somáticos más frecuentes: B, C1, C2, D y E, colocando en un portaobjetos una gota del antisuero y una gota del antígeno, para luego mezclar con un asa. Se rota el portaobjetos por un minuto y se lee la reacción 8. -Positivo: formación de grumos (aglutinación), antes de un minuto. Se analizaron 709 cepas de salmonella spp. 453 provenientes de coprocultivos y 256 de alimentos crudos o cocidos durante el periodo de juinio agosto 2006. Para determinar el fenotipo las cepas fueron aisladas e identificadas bioquimicamente, posteriormente serotipificadas y se determino su patron de resistencia a los antibioticos; ampicilina, cloranfenicol, estreptomicina, sulfonamida y tetraciclina (ACSSuT). El genotipo se determino mediante la tecnica electroforesis de campos pulsados en gel de agarosa (PFPG), empleando la enzima de restricción Xbal y el posterior analisis estadistico. De la serotipificacion de los 709 cepas se identificaron 106 pertenecientes al grupo somatico B, las cuales correspondieron al serotipo S. Typhimurium lo que en porcentajes representa al 14.9% de 709. Las cepas de otros serotipos fueron excluidas del estudio. La distribución geografica indico que los estados de Jalisco, Yucatán, Nuevo Leon, Guanajuato, Tlaxcala, Colima y Puebla son los que presentan mayor incidencia de este serovar de Salmonella. El fenotipo DT104 se obtuvo estudiando las 106 cepas de S. Typhimurium y 34 dieron el fenotipo d resistencia ACSSuT a los 5 antimicrobianos, de los cuales 18 provenian de coprocultivos y las 15 restantes de alimentos crudos o cocidos. La PFGE y el analisis bioestadística permitieron determinar que existen por lo menos 4 clonas diferentes del fenotipo DT104 de Salmonella enterica Subespecie enterica serotipo Typhimurium en Mexico según el patron de bandas obtenidas. En Mexico esta presente la cepa multiresistente DT104 de Salmonella enterica y existen diferentes clonas circulantes, por o que el riesgo potencial para la Salud Publica dbe advertirse, ya que no se habian realizado estudios de la presencia de este fenotipo en la Republica Mexicana. Impacto Deben adoptarse medidas sanitarias para prevenir la contaminación de alimentos y agua por roedores y otros animales que excretan Salmonella spp. La carne de aves,

33
bovinos y huevos deben ser cocinados adecuadamente. No se debe permitir que los portadores trabajen manejando alimentos, y es importante implementar programas de vigilancia con estrictas medidas higiénicas, así como caracterizar las tendencias de prevalencia y detectar brotes epidemiológicos con una comunicación rápida a las autoridades correspondientes 10,12. El Sistema Cooperativo de Vigilancia Internacional “Salm-Net” en Europa, facilita el intercambio de información para prevenir la infección humana asociada con productos agrícolas y alimentos manufacturados de amplia distribución universal 15. La detección y tratamiento de los portadores constituyen el mejor mecanismo de control de la fiebre tifoidea 12. En Europa ha sido creada Enter-net, red internacional para la vigilancia de infecciones gastrointestinales en el hombre. Esta red monitorea principalmente Salmonella spp y Escherichia coli O157 y uno de sus principales objetivos es establecer una base de datos internacional de casos de Salmonella spp con actualización periódica 38. Otra Red Europea Salm-Net, cuyo objetivo es prevenir las salmonelosis humanas en la Unión Europea mediante el refuerzo de la vigilancia internacional, con la creación de laboratorios y una base de datos disponible a todos. La red está compuesta por microbiólogos y epidemiólogos responsables de la vigilancia nacional de la salmonelosis en 14 países europeos. Además Salm-Net tiende a estandarizar los métodos de tipificación de fagos de S. Enteriditis, S. Typhimurium y S. Virchow 38. En una de las tesis se detectaron formas resitentes a los antibioticos denominadas cepas DT-104 que no habían sido detectadas en México y vale la pena vigilar y evitar que se diseminen y den brotes porque se comportarían de manera anómala y serian muy difícil de erradicar con el tratamiento antibiótico tradicional. CULTIVO, IDENTIFICACION Y ESTANDARIZACION DE LA REACCION EN CADENA DE LA POLIMERASA (PCR) PARA EL DIAGNOSTICO Mycoplasma pneumoniae. M. pneumoniae no puede ser diferenciada clínicamente de la neumonía causada por otras bacterias o virus.. Por lo que es importante realizar un diagnostico especifico, debido a que el tratamiento de M. pneumoniae con antibióticos beta lactámicos es inefectivo; mientras que el tratamiento con macrólidos o tetraciclinas pueden reducir marcadamente la evolución de la enfermedad Se subestima su etiología porque los síntomas son poco específicos, y que ademas, existen pocos laboratorios que ofrecen la búsqueda d esta bacteria , y carencia de métodos de diagnostico rápidos. Los laboratorios clínicos enfrentan varias dificultades para su detección, debido a que los metodos de cultivo son relativamente insensibles y requieren de hasta 3 semanas para su desarrollo de M. pneumoniae, Por lo que un metodo sensible y especifico para M. pneumoniae es mediante la técnica de PCR, que es el objetivo del presente trabajo y determinar la incidencia de esta en pacientes con infecciones respiratorias (IRA), tanto en niños como en adultos. Se llevo a cabo la estandarización de la técnica de PCR utilizando la cepa d referencia Eaton de M. pneumoniae proporcionada por el laboratorio d Bacteriología Médica de la Escuela Nacional de Ciencias Biologicas (ENCB), a la cual se le realizo la extracción del DNA, se le determino la pureza, integridad y concentración. Se realizaron curvas de MgCl2, de iniciadores MP5-1 y MP5-2, así como la determinación de la temperatura óptima de

34
alineación de estos, para llevar a cabo la amplificación de un fragmento de 104 pb, correspondiente a M. pneumoniae, la cual se utilizo para determinar la presencia de M. pneumoniae en 100 muestras clínicas proporcionadas por Unidad de Medicina No 2, de las cuales no se detecto la presencia de M. pneumoniae. Impacto. Mycoplasma pneumoniae es un patógeno común del tracto respiratorio humano. Se ha encontrado un promedio de infecciones en 10% al 20% de pacientes pediátricos hospitalizados y del 20% al 40% en pacientes pediátricos ambulatorios con neumonía, pero en México no se busca en los laboratorios de rutina aunque ya hay mucha metodología descrita y empleada en otros paidses aquí se debe de buscar. Resultado. Según las metas se cumplió el 100%
Enero 2007 - Diciembre 2007 Clave del Proyecto: 20070200
Expresión de factores de virulencia de Helicobacter pylori y Haemophilus influenzae, detección de micoplasmas y ureaplasmas por PCR y detección de cepas DT104 de Salmonella ssp.
1. Se seleccionaron y determino el tipo de muestra que se iba a estudiar de acuerdo con cada protocolo de tesis y se hicieron juntas en los hospitales correspondientes para obtener legalmente estas cepas de acuerdo con los comités de ética y proyectos registrados en esas instituciones para poder hacer los estudios correspondientes en cada caso, obtener las colecciones correspondientes. Enseguida se procedió a dar entrenamiento a los alumnos sobre la conservación de cepas y se organizo un Simposio sobre conservación de cepas que tuvo mucha demanda y sirvió para concientizar al grupo sobre la importancia de tener congeladores que sirvan, planta de luz y servicio de nitrógeno líquido, sino todo se pierde. 2. Se hicieron los ensayos correspondientes de reacción en cadena de polimerasa PCR para diferentes géneros de acuerdo con los protocolos establecidos y se concreto en una tesis de Maestría los resultados acerca de los genes glmM ureA cagA, bab A2 ,vac A en la obtención de cepas virulentas triple positivas, se estandarizó la prueba con cepas de referencia y cepas clínicas y se titularon cuatro alumnos cada uno aporto resultados en su tesis que permiten plantear nuevas hipótesis y metas para 2008 y servirán para implementar las publicaciones del grupo 3. El modelo en cultivo celular de células Hep 2 resulto muy difícil de implementar, ya que algunas veces hubo contaminación cruzada, falta de reactivos como suero fetal de ternera certificado que cada vez es más caro, o bien cuando se infectaba con Helicobacter pylori daba resultados pobres, por lo menos se logro estandarizar y confirmar que, efectivamente cuando hay infección por Helicobacter pylori se induce la producción de metaloprotesas mmp-9 y no hay producción de metaloproteasas mmp-2, este descubrimiento es importante para la patogenicidad porque plantea la liberación de mediadores de inflamación independientes de la respuesta inmune como la IL-8 4. Se seleccionó un grupo de cepas de Haemophilus influenzae que están en diferentes serogrupos para estudiar si tienen factores de virulencia, buscando los genes de la isla de patogenicidad mediante pruebas PCR ICEHin 1056, de acuerdo con la meta número 5, se trabajaron para obtener sobrenadantes en cultivo en caldo BHI suplementado, con ese extracto crudo filtrado se hicieron ensayos en cultivo celular Hep-2 para medir el efecto de

35
adherencia con bacterias completas y efecto citotóxico con los extractos, estos resultados fueron presentado en varios congresos como trabajos libres. 5 Se busco el gen hap con pruebas de PCR, y se midió su expresión en cultivos celulares con bacterias completas de las hap positivas estaban presentes en todas las cepas de Haemophilus influenzae y daban efecto citotóxico y adherencia que fue dificil de clasificar porque hubo patrones localizados, agregativos y difusos. Se presento una tesis de licenciatura. 6 Se estandarizó la prueba de PCR para identificación de Mycoplasma pneumoniae con una cepa de referencia, se presentó un cartel y una tesis de licenciatura con los resultados obtenidos. Se plantea la dificultad de trabajar con muestras de expectoración, ya que la purificación del DNA es difícil y puede dar falsas negativas, motivo por el cual se implementó una técnica especial para obtener DNA de mejor calidad, pero para poderlo publicar es necesario trabajar con un número mayor de muestras. Ureaplasma urealyticum necesita muestras adecuadas y una mejor estandarización, al igual que con las expectoraciones, si se trabaja exudados vaginales debe obtenerse DNA de buena calidad 7. Se trabajaron cepas de Salmonella typhimurium que no se sabia su patrón de susceptibilidad a los antibióticos y con ellas se pudo confirmar que es posible que en México estén circulado cepas del fenotipo DT-104 que son importantes desde el punto de vista epidemiológico. Se hizo electroforesis de campo pulsado PFGE para entender las relaciones genotípicas de los aislados. Se tituló un alumno de licenciatura
5. Impacto.
El beneficio social directo es sobre la formación de recursos humanos de alto nivel en el área de Salud, el beneficio social es que las personas formadas bajo este proyecto tienen mejor criterio y pueden defenderse en el campo laboral relacionado con diagnóstico microbiológico, investigación o pueden preparar a otros grupos de profesionales en el trabajo.
En este proyecto se logro un impacto elevado en la formación de recursos humanos para el Sector Social y Económico aunque se abordaron líneas de investigación aparentemente diversas, los resultaron fueron positivos en Helicobacter pylori hubo 5 tesis de licenciatura, dos de maestría; dos de licenciatura de Enterococcus spp, en Haemophilus influenzae hubo 2 tesis de licenciatura y en Mycoplasma una, con buenas aportaciones metodológicas y en Salmonella 2 tesis de licenciatura el trabajo experimental en una de ellas seria para una publicación. Las dos tesis de doctorado tienen importancia en el estudio de la relación Huésped Parásito y en el estudio de patogénesis de las enfermedades infecciosas. En resistencia antibióticos se abordo en el tema en casi todas las bacteria estudiadas hubo una publicación en la revista latinomericana de microbiología que resume bien la importancia e impacto de estos estudios no solo para el paciente, sino para el médico que usualmente solo recurre al diagnóstico anatomopatológico pero estamos demostrando que se pueden obtener resultados de alto impacto con técnicas modernas de biología molecular y para bacterias fastidiosas como Helicobacter pylori o Mycoplasma incluyendo otras oportunistas como el género Aeromonas que son muy importantes al igual que

36
Salmonella por su participación en diarrea persistente y en brotes de ETA(Enfermedades trasmitidas por los alimentos obtenidas a partir de productos diversos, aquí hay que considerar la dificultad para obtener muestras adecuadas para diagnóstico, ya que los alumnos y los directores de tesis nos tenemos que apegar a los lineamientos de ética de los hospitales o clínicas y también dentro del propio laboratorio debemos de conservar los reglamentos en cuanto al manejo de productos biológico infecciosos y enseñar además de las técnicas usuales de microbiología las apegada al protocolo de investigación, medidas de asepsia y higiene y control de calidad, motivo por el cual en forma conjunta se insistió en la importancia de la conservación de cepas. Evitar las contaminaciones cruzadas en cultivos celulares y en pruebas de biología molecular. Toda esta formación profesional repercute en la madurez y desempeño que tienen los egresados con esta formación a nivel profesional y de investigación.
ABREVIATURAS
ABREVIATURA SIGNIFICADO
AINE Fármacos antiinflamatorios no esteriodeos
ATCC American Type Culture Collection, USA
(Colección Americana de Cultivos Tipo)
cag- PAI Isla de patogenicidad de cag
CagA
cagA
cagE
Proteína A asociada a citotoxicidad de H. Pylori
Gen que codifica la proteína asociada a citotoxina A
Gen que codifica para el sistema de secreción tipo IV en
H. pylori
CIM Concentración Inhibitoria Mínima
CLSI Clinical and Laboratory Standards Institute (Comite Nacional de Estándares de Laboratorios Clínicos)
DAR Dolor abdominal recurrente
EAP Enfermedad ácido péptica
ERGE Enfermedad por reflujo gastroesofágico
FDA Food and Drug Administration, USA

37
(Administración de drogas y alimentos)
flaA
flan
Genes que codifican para la movilidad de H. pylori
glmM Gen que codifica para una fosfoglucosamina mutasa de
H. pylori.
HIMFG Hospital Infantil de México Federico Gómez
IgA
IgG
IgM
Inmunoglobulina A -secretora
Inmunoglobulina G -memoria
Inmunoglobulina M –reconocimiento
IL-8 Interleucina -8
MALT Linfoma de tejido linfoide asociado a mucosas
PCR Reacción en cadena de la polimerasa
( R ) Cepa resistente
rRNA 16S y 23S Ácido ribonucleico ribosomal fracción 16S y 23S
( S ) Cepa sensible
UBT Urea Breath Test, Prueba de aliento
VacA
vacA
Proteína A citotoxina vacuolizante de H. pylori
Gen que codifica para la citotoxina vacuolizante A